м„ңлЎ
1. м—°кө¬мқҳ н•„мҡ”м„ұ
비л§ҢмқҖ к°Ғмў… м„ұмқёлі‘мқ„ мң л°ңн•ҳлҠ” вҖҳм§Ҳлі‘вҖҷмңјлЎң м„ёкі„ліҙкұҙкё°кө¬(WHO)к°Җ к·ңм •н• м •лҸ„лЎң м „ м„ёкі„м Ғ л¬ём ңлЎң мЈјлӘ©лҗҳкі мһҲлӢӨ. 비л§Ңмқҳ мӣҗмқёмқҖ м—¬лҹ¬ мҡ”мқёмқҙ ліөн•©м ҒмңјлЎң мһ‘мҡ©н•ңлӢӨ. кё°ліём ҒмңјлЎң м—җл„Ҳм§Җ м„ӯм·ЁмҷҖ мҶҢ비мқҳ л¶Ҳк· нҳ•мңјлЎң м§Җл°©мқҙ 축м Ғлҗҳкі , л¶Җк°Җм ҒмңјлЎң л°”мҒң нҳ„лҢҖмқёл“Өмқҳ л¶Ҳк·ңм№ҷм Ғмқё мӢқмҠөкҙҖ, мһҗлҸҷнҷ”лЎң мқён•ң мҡҙлҸҷл¶ҖмЎұ, мҠӨнҠёл ҲмҠӨ к°ҷмқҖ м •мӢ м Ғ л¬ём ң к·ёлҰ¬кі мқјл¶Җ мң м „м Ғмқё мӣҗмқё л“ұмқҙ мҳҒн–Ҙмқ„ лҜём№ңлӢӨ. мқҙмҷҖ н•Ёк»ҳ лҳҗ н•ҳлӮҳмқҳ мҡ”мқёмқҙ 비л§Ңмқҳ мӣҗмқёмңјлЎң мЈјлӘ©мқ„ л°ӣкі мһҲлӢӨ. мқҙ мҡ”мқёмқҖ мқёк°„мқ„ нҸ¬н•Ён•ң лҸҷл¬јкіј мӢқл¬јм—җм„ң 24мӢңк°„мқ„ мЈјкё°лЎң н•ҳлҠ” мғқмІҙлҰ¬л“¬(circadian rhythm)мңјлЎң, мІҙмҳЁмЎ°м Ҳ, нҳёлҘҙлӘ¬ мғқм„ұ, мҲҳл©ҙ, м„ӯмӢқ л“ұкіј к°ҷмқҖ мғқл¬јн•ҷм Ғ, мғқлҰ¬н•ҷм Ғ, н–үлҸҷн•ҷм Ғ нҷңлҸҷм—җ кҙҖм—¬н•ңлӢӨ[1, 2].
мғқмІҙлҰ¬л“¬мқҖ мқјл Ёмқҳ мғқмІҙмӢңкі„ мң м „мһҗмқҳ мЎ°м Ҳм—җ мқҳн•ҳм—¬ мқҙлЈЁм–ҙ진лӢӨ. мӮ¬лһҢмқҳ кІҪмҡ°, circadian locomotor output cycles kaput (CLOCK), brain and muscle ARNT-like protein 1 (BMAL1), period 1-3 (PER1-3), Cryptochrome 1-2 (CRY 1-2), reverse erythroblastosis virus (REV-ERBОұ), к·ёлҰ¬кі retinoid-related orphan receptor- (RORОұ)мҷҖ к°ҷмқҖ мғқмІҙмӢңкі„мқҳ мң м „мһҗл“Өмқҙ мҶҢлҮҢ, мӨ‘лҮҢ, мӢңмғҒн•ҳл¶ҖмҷҖ к°ҷмқҖ лҮҢмЎ°м§Ғ лҝҗ м•„лӢҲлқј мӢ¬мһҘ, нҸҗ, м§Җл°©, нҳҲкҙҖ, мӢ мһҘкіј к°ҷмқҖ мһҘкё°л“Өм—җлҸ„ мЎҙмһ¬н•ңлӢӨ. лҮҢ мӢңмғҒ н•ҳл¶Җмқҳ мӢңкөҗм°ЁмғҒн•ө(Suprachiasmatic Nucleus, SCN)м—җ мЎҙмһ¬н•ҳлҠ” мӢ кІҪм„ёнҸ¬м—җ мЎҙмһ¬н•ҳлҠ” мғқмІҙмӢңкі„ мң м „мһҗл“ӨмқҖ к°Ғ мЎ°м§Ғм„ёнҸ¬мқҳ кі мң н•ң лҰ¬л“¬мқ„ лҸҷкё°нҷ”(synchronization)мӢңмјң к°ңмІҙмқҳ н–үлҸҷм–‘мғҒмқ„ мЎ°м Ҳн•ҳлҠ” мӨ‘추мӢңкі„(central Clock)мқҳ кё°лҠҘмқ„ н•ҳлҠ” л°ҳл©ҙ, к°Ғ мЎ°м§Ғм—җ мЎҙмһ¬н•ҳлҠ” мң м „мһҗл“ӨмқҖ л§җмҙҲмӢңкі„(peripheral Clock)лЎң мһ‘мҡ©н•ҳм—¬ нҳёлҘҙлӘ¬кіј мӢ кІҪм „лӢ¬л¬јм§Ҳмқ„ нҶөн•ҙ мһҘкё°мқҳ нҷңлҸҷмқ„ мӢңк°„мқҳ нқҗлҰ„м—җ л”°лқј мЎ°м Ҳн•ҳлҠ” кІғмқҙ л°қнҳҖмЎҢлӢӨ[3].
мқҙ мғқмІҙмӢңкі„мқҳ көҗлһҖм—җ мқҳн•ҙ 비л§Ңкіј мӢ 진лҢҖмӮ¬мқҳ мһҘм• к°Җ мқјм–ҙлӮҳлҠ” лҢҖмӮ¬мҰқнӣ„кө°мқҳ мң„н—ҳмқ„ мҰқк°ҖмӢңнӮЁлӢӨлҠ” мӮ¬мӢӨмқҙ мһ…мҰқлҗҳм—ҲлӢӨ. Turek (2005)мқҳ м—°кө¬м—җ мқҳн•ҳл©ҙ, мғқмІҙмӢңкі„ мң м „мһҗ мӨ‘ н•ҳлӮҳмқё Clockм—җ лҸҢм—°ліҖмқҙк°Җ мқјм–ҙлӮң мғқмҘҗл“Өм—җкІҢ 24мӢңк°„ мғқмІҙмЈјкё°мқҳ көҗлһҖмқҙ мқјм–ҙлӮҳл©ҙм„ң мІҙмӨ‘кіј лЁ№мқҙмқҳ м„ӯм·Ёк°Җ мҰқк°Җн•ҳкі нҳҲлӢ№ л°Ҹ нҳҲмӨ‘ мҪңл ҲмҠӨн…ҢлЎӨ(cholesterol)кіј нҠёлҰ¬кёҖлҰ¬м„ёлҰ¬л“ң(triglycerides, TG)к°Җ мҰқк°Җн•ҳлҠ” лҢҖмӮ¬мҰқнӣ„кө°мқҳ мҰқмғҒмқҙ мқјм–ҙлӮ¬мқҢмқ„ ліҙкі н•ҳмҳҖлӢӨ[4]. н•ңнҺё, мң м „мһҗ Periodмқҳ л°ңнҳ„мқҙ м–өм ңлҗң мғқмҘҗ(period knockout mice)лҘј лӮ®кіј л°Өмқҳ мЈјкё°к°Җ 12мӢңк°„(м •мғҒ), 4мӢңк°„ лҳҗлҠ” 18мӢңк°„ к°„кІ©мңјлЎң мЎ°м •лҗң м„ё к°Җм§Җмқҳ мЎ°кұҙм—җм„ң мӮ¬мңЎн•ҳмҳҖмқ„ л•Ң м •мғҒмғқмҘҗм—җ 비н•ҳм—¬ н–үлҸҷл Ҙмқҙ к°җнҮҙлҗҳкі м„ӯмӢқмқҙ мҰқк°ҖлҗҳлҠ” кІғмқ„ кҙҖм°°н•ҳмҳҖлӢӨ[5]. мқҙлҠ” лӮ®кіј л°Өмқҳ мЈјкё°к°Җ мғқмІҙмӢңкі„ мң м „мһҗмҷҖ мғҒнҳёмһ‘мҡ©мқҙ мқјм–ҙлӮҳл©° мғқмІҙмӢңкі„мқҳ көҗлһҖмқҙ мғқлҰ¬м Ғ, лі‘лҰ¬м Ғ ліҖнҷ”лҘј мқјмңјнӮ¬ мҲҳ мһҲмқҢмқ„ ліҙм—¬мЈјлҠ” кІ°кіјмқҙлӢӨ. нҳ„лҢҖм—җлҠ” мғқнҷңмҠөкҙҖкіј нҷҳкІҪмқҳ мҳҒн–ҘмңјлЎң мқјмЈјкё°лҰ¬л“¬мқҳ көҗлһҖмқҙ л°ңмғқн•ҳлҠ” мҳҲк°Җ л§Һм•„м§Җкі мһҲлӢӨ. нҠ№нһҲ к№Ём–ҙмһҲлҠ” мӢңк°„мқҙ кёём–ҙм§Җкі көҗлҢҖк·јл¬ҙмһҗл“Өкіј к°ҷмқҙ лӮ®кіј л°Өмқҙ л’Өл°”лҖҢлҠ” нҳ„мғҒмқҙ л№ҲлІҲн•ҳкІҢ мқјм–ҙлӮҳкі мһҲм–ҙ мқҙлҹ¬н•ң мӮ¬лһҢл“ӨмқҖ мҲҳл©ҙл¶ҖмЎұкіј н”јлЎң, нҳёлҘҙлӘ¬мқҙ м—°кҙҖлҗң м§Ҳнҷҳм—җ мқҙнҷҳлҗҳлҠ” нҷ•лҘ мқҙ лҶ’м•„м§Җкі мһҲлӢӨ[6]. лҝҗл§Ң м•„лӢҲлқј к°Ғ мЎ°м§Ғм—җм„ңлҠ” л§җмҙҲ мғқмІҙмӢңкі„ мң м „мһҗмқҳ л°ңнҳ„мқҙ 붕кҙҙлҗҳкі м§Җл°©м„ёнҸ¬мқҳ м§Җл°©лҢҖмӮ¬, м·ҢмһҘмқҳ мқёмҠҗлҰ° 분비, к°„кіј к·јмңЎмқҳ лӢ№лҢҖмӮ¬, мһҘлӮҙ мҶҢнҷ”нқЎмҲҳ л°Ҹ л©ҙм—ӯм„ёнҸ¬мқҳ кё°лҠҘмқ„ мғҒмӢӨн•ҳм—¬ кІ°көӯ м—җл„Ҳм§Җ н•ӯмғҒм„ұмқҳ көҗлһҖмқҙ мқјм–ҙлӮҳкі лҢҖмӮ¬м„ұ м§Ҳнҷҳмқ„ мқјмңјнӮӨлҠ” кІ°кіјк°Җ л°ңмғқн•ҳлҠ” мӮ¬лЎҖл“Өмқҙ ліҙкі лҗҳкі мһҲлӢӨ[7]. лҳҗн•ң мғқмІҙмӢңкі„мқҳ көҗлһҖмқҖ мҲҳлӘ…мқ„ лӢЁм¶•мӢңнӮӨлҠ” кІ°кіјлҸ„ к°Җм ёмҳӨкё° л•Ңл¬ём—җ[5] м Ғм Ҳн•ң лӮ®кіј л°Өмқҳ мЈјкё°м—җ л”°лҘё мғқмІҙмӢңкі„мқҳ мЎ°м Ҳмқҙ м •мғҒм Ғмқё мғқлҰ¬нҳ„мғҒмқ„ мң м§Җн•ҳкі м§Ҳлі‘мқҳ л°ңмғқмқ„ к°җмҶҢмӢңнӮ¬ мҲҳ мһҲлҠ” мӨ‘мҡ”н•ң мҡ”мқёмқҙлқј н•ҳкІ лӢӨ.
нҳ„лҢҖмӮ¬нҡҢм—җм„ң м җм°Ё мҰқк°Җн•ҳлҠ” мҠӨнҠёл ҲмҠӨлҠ” 비л§Ңмқ„ мң л°ңн•ҳлҠ” мҡ”мқёмңјлЎң м§ҖлӘ©лҗҳм—ҲлӢӨ. мҠӨнҠёл ҲмҠӨм—җ л…ём¶ңлҗҳл©ҙ н•ӯмғҒм„ұмқ„ мң м§Җн•ҳкё° мң„н•ҙ көҗк°җ мӢ кІҪм—җ мқҳн•ң мһҗмңЁмӢ кІҪкі„ л°ҳмқ‘кіј мӢңмғҒн•ҳл¶Җ-лҮҢн•ҳмҲҳмІҙ-л¶ҖмӢ (HPA axis)м—җ мқҳн•ң мӨ‘추мӢ кІҪкі„ л°ҳмқ‘мқҙ мқјм–ҙлӮҳкі мӢ мІҙмқҳ м—¬лҹ¬ кё°кҙҖм—җм„ң нҠ№мқҙм Ғ м Ғмқ‘ л°ҳмқ‘л“Өмқҙ нҷңм„ұнҷ”лҗңлӢӨ. мқҙлҹ° л°ҳмқ‘мқҖ м—җн”јл„Өн”„лҰ°(epinephrine), л…ём–ҙм—җн”јл„Өн”„лҰ°(norepinephrine) к·ёлҰ¬кі мҪ”нӢ°мҶ”(cortisol)мқҳ 분비лҘј мҙү진мӢңнӮЁлӢӨ. көҗк°җмӢ кІҪкі„м—җм„ң 분비лҗҳлҠ” м№ҙн…ҢмҪңлқјлҜј(catecholamine)мқҖ лҢҖн•ӯнҡҢн”јл°ҳмқ‘(fight-or-flight response)мқ„ мң лҸ„н•ҳл©°, мқҙл•Ң мӢ¬л°•лҸҷ мҰқк°Җ, нҳҲм••мғҒмҠ№кіј к°ҷмқҖ мӢ мІҙм Ғ ліҖнҷ”к°Җ мқјм–ҙлӮңлӢӨ. л§Ңм„ұ мҠӨнҠёл ҲмҠӨм—җ л…ём¶ңлҗҳм–ҙ мҪ”нӢ°мҶ” к°ҷмқҖ кёҖлЈЁмҪ”мҪ”лҘҙнӢ°мҪ”мқҙл“ң(glucocorticoid)к°Җ 과분비лҗҳл©ҙ лҢҖмӮ¬кіјм • мқҙмғҒ, мҶҢнҷ”кё°кі„ мқҙмғҒ, л©ҙм—ӯн•ҷм Ғ мқҙмғҒ, м •мӢ м Ғмқё мһҘм• л“ұмқҙ л°ңмғқн•ҳкі , мқҙлЎң мқён•ҙ лҢҖмӮ¬мҰқнӣ„кө°, кі нҳҲм••, мӢ¬мһҘм§Ҳнҷҳ л“ұмқҳ м§Ҳлі‘мқҙ мң л°ңлҗ к°ҖлҠҘм„ұмқҙ лҶ’아진лӢӨ[8].
мғқмҘҗм—җкІҢ кі м§Җл°©мӢқмқҙлҘј кіөкёүн•ҳм—¬ 비л§Ң л°ңмғқмқ„ мң лҸ„н•ҳлҠ” мӢӨн—ҳ лӘЁлҚёмқҖ 비л§Ң, лӢ№лҮЁ, кі м§ҖнҳҲмҰқкіј лҢҖмӮ¬мҰқнӣ„кө°мқ„ м—°кө¬н•ҳкё° мң„н•ҳм—¬ мһҗмЈј мқҙмҡ©лҗңлӢӨ[9]. мғқмҘҗм—җкІҢ 45% кі м§Җл°©мӢқмқҙлҘј кіөкёүн•ҳлҠ” кІҪмҡ° 16мЈј мқҙнӣ„л¶Җн„°, 60% кі м§Җл°©мӢқмқҙлҘј кіөкёүн•ҳлҠ” кІҪмҡ°м—җлҠ” 8мЈј мқҙнӣ„л¶Җн„° нҳҲмӨ‘ нҸ¬лҸ„лӢ№, м§Җм§Ҳ м§Җн‘ңмҷҖ н•Ёк»ҳ мқёмҠҗлҰ° м Җн•ӯм„ұмқҙ мқјм–ҙлӮңлӢӨ. ліё м—°кө¬лҠ” кі м§Җл°© н•Ёмң лҹүмқҙ лӢӨлҘё мӢқмқҙлҘј мЈјм–ҙ мғқмҘҗм—җкІҢ мһҘ, лӢЁкё°м ҒмңјлЎң лӮҳнғҖлӮҳлҠ” мІҙмӨ‘мқҳ ліҖнҷ”, м„ӯмӢқ, 비л§Ң л°Ҹ лҢҖмӮ¬мҰқнӣ„кө°кіј м—°кҙҖлҗң мң м „мһҗмқҳ ліҖнҷ”мҷҖ н•Ёк»ҳ мғқмІҙмӢңкі„ мң м „мһҗл“Өмқҙ 비л§Ңм—җ мқҳн•ҙ м–ҙл–»кІҢ ліҖнҷ”н•ҳлҠ”м§Җ мӮҙнҺҙліҙкі кі м§Җл°©мӢқмқҙлҘј м„ӯм·Ён•ң мғқмҘҗм—җкІҢ л¶ҖлҸҷ мҠӨнҠёл ҲмҠӨлҘј к°Җн•ҳм—¬ 비л§Ңмқҳ 진н–үмқҙ к°ҖмҶҚнҷ” лҗ мҲҳ мһҲлҠ”м§Җ, к·ёлҰ¬кі 비л§Ңмқҳ 진н–үмқҙ лҢҖмӮ¬кҙҖл Ё мң м „мһҗмҷҖ мғқмІҙмӢңкі„ мң м „мһҗмқҳ ліҖнҷ”лҘј мң лҸ„н•ҳлҠ”м§Җ м•Ңм•„ліҙкі мһҗ н•ҳмҳҖлӢӨ.
2. м—°кө¬ лӘ©м Ғ
ліё м—°кө¬лҠ” мғқмҘҗ лҸҷл¬јлӘЁлҚём—җ кі м§Җл°©мӢқмқҙлҘј кіөкёүн•ҳм—¬ 비л§Ңмқ„ мң лҸ„н•ң нӣ„ 비л§Ңмқҙ л°ңмғқн•ҳлҠ” кіјм •м—җм„ң мӢқмқҙ лӮҙ кі м§Җл°© н•Ёмң мңЁмқҳ м°Ёмқҙм—җ л”°лҘё мң м „мһҗмқҳ л°ңнҳ„, к·ёлҰ¬кі мҠӨнҠёл ҲмҠӨ м Ғмҡ©мқ„ н•ң кІҪмҡ° ліҖнҷ”н•ҳлҠ” мң м „мһҗ л°ңнҳ„мқ„ нҷ•мқён•ҳкі мһҗ н•ҳмҳҖлӢӨ. кө¬мІҙм Ғ лӘ©м ҒмқҖ лӢӨмқҢкіј к°ҷлӢӨ.
1) 45%мҷҖ 60%мқҳ м§Җл°©н•Ёмң мңЁмқҙ лӢӨлҘё мӢқмқҙлҘј кіөкёүн•ң мғқмҘҗмқҳ мІҙмӨ‘, нҳҲлӢ№, нҳҲмІӯм§Җм§Ҳ, мӢқмқҙнҡЁмңЁ л“ұ мғқлҰ¬н•ҷм Ғ ліҖнҷ”лҘј мёЎм •н•ңлӢӨ.
2) 비л§Ңмқҙ л°ңмғқн•ҳлҠ” кіјм •м—җм„ң м§Җл°©н•Ёмң мңЁмқҳ м°Ёмқҙм—җ л”°лҘё лҢҖмӮ¬кҙҖл Ё мң м „мһҗмқҳ ліҖнҷ”лҘј нҢҢм•…н•ңлӢӨ.
3) 비л§Ңмқҙ л°ңмғқн•ҳлҠ” кіјм •м—җм„ң л§җмҙҲ мғқмІҙмӢңкі„ мң м „мһҗмқҳ ліҖнҷ”лҘј нҢҢм•…н•ңлӢӨ.
4) мҠӨнҠёл ҲмҠӨмқҳ м Ғмҡ©мқҙ 비л§Ңмқ„ к°ҖмҶҚнҷ”н•ҳлҠ”м§Җ мӮҙнҺҙліҙкі лҢҖмӮ¬кҙҖл Ё мң м „мһҗмҷҖ л§җмҙҲ мғқмІҙмӢңкі„ мң м „мһҗмқҳ ліҖнҷ”лҘј мң лҸ„н•ҳлҠ”м§Җ нҢҢм•…н•ңлӢӨ.
м—°кө¬ л°©лІ•
1. м—°кө¬м„Өкі„
ліё м—°кө¬лҠ” мғқмҘҗ лҸҷл¬јлӘЁлҚёмқ„ мқҙмҡ©н•ҳм—¬ кі м§Җл°©мӢқмқҙмқҳ м§Җл°© н•Ёмң мңЁм—җ л”°лҘё мғқлҰ¬н•ҷм Ғ ліҖнҷ”мҷҖ лҢҖмӮ¬кҙҖл Ё мң м „мһҗ л°ңнҳ„ м •лҸ„, к·ёлҰ¬кі л§җмҙҲ мғқмІҙмӢңкі„ л°ңнҳ„мқҳ ліҖнҷ”лҘј мёЎм •н•ң мӢӨн—ҳм—°кө¬мқҙлӢӨ. лҳҗн•ң мҠӨнҠёл ҲмҠӨ л…ём¶ңм—җ мқҳн•ҳм—¬ 비л§Ң м •лҸ„мқҳ ліҖнҷ”лҘј м•Ңм•„ліё мӢӨн—ҳм—°кө¬мқҙлӢӨ.
2. м—°кө¬лҢҖмғҒ
мӢӨн—ҳлҸҷл¬јмқҖ 6мЈјл №мқҳ C57BL/6J (мӨ‘м•ҷмӢӨн—ҳлҸҷл¬ј, Korea) л§Ҳмҡ°мҠӨлҘј 1мЈјмқјк°„ мқјл°ҳ кі нҳ•мӮ¬лЈҢлЎң мӮ¬мңЎн•ҳл©ҙм„ң нҷҳкІҪм—җ м Ғмқ‘мӢңмј°лӢӨ. мӢӨн—ҳлҸҷл¬јмқҖ л¬јкіј мӮ¬лЈҢлҘј 충분нһҲ кіөкёүн•ҳмҳҖмңјл©°(ad libitum), мӮ¬мңЎмЎ°кұҙмқҖ мҳЁлҸ„ 23Вұ1В°C, мҠөлҸ„ 50Вұ5%, лӮ®кіј л°Өмқҳ мЈјкё°лҠ” 12мӢңк°„мңјлЎң мЎ°м Ҳн•ҳмҳҖлӢӨ. 1мЈјмқјк°„мқҳ м Ғмқ‘кё°к°„мқ„ кұ°м№ң мғқмҘҗлҠ” кё°ліё мІҙмӨ‘мқ„ мёЎм •н•ң нӣ„ л¬ҙмһ‘мң„лЎң к°Ғ мӢӨн—ҳкө°м—җ л°°м •н•ҳмҳҖлӢӨ. м§Җл°©н•Ёмң лҹүмқҳ м°Ёмқҙм—җ л”°лҘё мӢӨн—ҳмқҖ 8мЈј мӮ¬мңЎн•ҳмҳҖмңјл©°, мҠӨнҠёл ҲмҠӨ м Ғмҡ©мқ„ мң„н•ң мӢӨн—ҳмқҖ 16мЈјк°„ мӮ¬мңЎн•ҳлҠ” мҙқ м—¬м„Ҝ к·ёлЈ№мңјлЎң лӮҳлҲ„м—Ҳкі , нғҲлқҪмңЁмқ„ кі л Өн•ҳм—¬ кө° лӢ№ 5л§ҲлҰ¬ мқҙмғҒмқ„ нҷ•ліҙн•ҳмҳҖлӢӨ. мӢӨн—ҳмқҳ м „ кіјм •мқҖ YлҢҖн•ҷкөҗ лҸҷл¬јмңӨлҰ¬мң„мӣҗнҡҢмқҳ мҠ№мқё(2013-0233)мқ„ л°ӣмқҖ нӣ„ к·ңм •м—җ л”°лқј мӢӨмӢңн•ҳмҳҖлӢӨ.
3. мӢӨн—ҳл°©лІ•
1) кі м§Җл°©мӢқмқҙмқҳ кіөкёү
мӢӨн—ҳлҸҷл¬јмқҖ мІҙмӨ‘ мёЎм •мқ„ н•ң нӣ„, л¬ҙмһ‘мң„лЎң в‘ м •мғҒмӢқмқҙ(Chow diet, CD), в‘Ў 45% м§Җл°©мқ„ н•Ёмң н•ң кі м§Җл°©мӢқмқҙ(45% high fat diet, 45HFD), в‘ў 60% м§Җл°©мқ„ н•Ёмң н•ң кі м§Җл°©мӢқмқҙ(60% high fat diet, 60HFD)лҘј кіөкёүл°ӣлҠ” м„ё кө°мңјлЎң 분лҘҳн•ҳм—¬ мҙқ 8мЈјк°„ мӮ¬мңЎн•ҳмҳҖлӢӨ.
2) мҠӨнҠёл ҲмҠӨмқҳ м Ғмҡ©
мӢӨн—ҳлҸҷл¬јмқҳ мҠӨнҠёл ҲмҠӨ лӘЁлҚёмқҖ л¶ҖлҸҷ мҠӨнҠёл ҲмҠӨ(immobilization stress)лҘј мӢӨмӢңн•ҳмҳҖлӢӨ. 3 cmГ—6 cm к°„кІ©мңјлЎң 6м№ёмқҳ м•„нҒ¬лҰҙ мјҖмқҙмҠӨлҘј м ңмһ‘н•ң нӣ„ лҡңк»‘м—җ нҶөн’Қмқҙ мһҳ лҗҳлҸ„лЎқ мһ‘мқҖ кө¬л©Қмқ„ м—¬лҹ¬ к°ң лҡ«м–ҙмЈјм—ҲлӢӨ[10]. мҠӨнҠёл ҲмҠӨ м Ғмҡ©мқҖ 45HFDкө°м—җкІҢ 16мЈј лҸҷм•Ҳ л§Өмқј мҳӨм „ 10мӢңм—җ 2мӢңк°„м”© к°Ғ мјҖмқҙмҠӨм—җ л„ЈлҠ” л°©лІ•мңјлЎң мӢңн–үн•ҳмҳҖмңјл©°(45HFD+St), лҢҖмЎ°кө°мқё CD, 45HFDлҠ” л¬јкіј лЁ№мқҙл§Ң л№јкі мһҗмң лЎӯкІҢ л‘җм—ҲлӢӨ.
3) мІҙмӨ‘ л°Ҹ мӢқмқҙнҡЁмңЁ мёЎм •
мӢӨн—ҳмӢқмқҙлҘј кіөкёүн•ң нӣ„ 1мЈјмқј к°„кІ©мңјлЎң л§Ө нҡҢ мқјм •н•ң мӢңк°„м—җ лҸҷл¬јмҡ© мІҙмӨ‘кі„(KC-500, CAS science, Daejeon, Korea)лЎң мёЎм •н•ҳмҳҖлӢӨ. кіөліөнҳҲлӢ№ мёЎм •мқ„ мң„н•ҳм—¬ м „лӮ м Җл…Ғл¶Җн„° 16мӢңк°„лҸҷм•Ҳ кёҲмӢқмӢңнӮЁ нӣ„ лӢӨмқҢ лӮ мҳӨм „м—җ кј¬лҰ¬л§җмҙҲнҳҲкҙҖмңјлЎңл¶Җн„° мҶҢлҹүмқҳ нҳҲм•Ўмқ„ мұ„м·Ён•ҳм—¬ ACCU-CHEK performa (Roche, Mannheim, Germany)мңјлЎң мёЎм •н•ҳмҳҖлӢӨ. мӢқмқҙлҠ” л§ӨмЈј лҸҷмқјн•ң мӢңк°„м—җ кіөкёүн•ҳмҳҖкі лӮЁмқҖ мӮ¬лЈҢмқҳ м–‘кіј к·ёлӮ кіөкёүн•ң мӮ¬лЈҢмқҳ м–‘мқ„ л§ӨмЈј мёЎм •н•ҳм—¬ к·ё м°ЁмқҙлЎң м„ӯм·Ён•ң мӢқмқҙлҹүмқ„ кі„мӮ° н•ҳмҳҖлӢӨ. мӢқмқҙнҡЁмңЁ(food efficiency ratio, FER)мқҖ мӢӨн—ҳ кё°к°„м—җ мҰқк°Җн•ң мІҙмӨ‘ мҰқк°Җлҹүмқ„ к°ҷмқҖ кё°к°„ лҸҷм•Ҳм—җ м„ӯм·Ён•ң мҙқмӢқмқҙлҹүмңјлЎң лӮҳлҲ„м–ҙ мӮ°м¶ңн•ҳмҳҖкі мӢқмқҙнҡЁмңЁ мӮ°м¶ңкіөмӢқмқҖ м•„лһҳмҷҖ к°ҷлӢӨ[11].
4) мұ„нҳҲ л°Ҹ мһҘкё° м Ғм¶ң
мӢӨн—ҳлҸҷл¬јмқҖ 8мЈј нӣ„мҷҖ 16мЈј нӣ„, 16мӢңк°„ кёҲмӢқмӢңнӮЁ нӣ„ Zoletil50 (VirbacKorea, Seoul, Korea) (30 mg/kg)кіј Rompun (BayerKorea, Seoul, Korea) (10 mg/kg)мқ„ ліөк°• мЈјмӮ¬н•ҳм—¬ л§Ҳм·Ё нӣ„ к°ңліөн•ҳмҳҖлӢӨ. ліөл¶ҖлҘј к°ңліөн•ң нӣ„ л©ёк· мЈјмӮ¬кё°лҘј мӮ¬мҡ©н•ҳм—¬ мӢ¬мһҘм—җм„ң нҳҲм•Ўмқ„ мұ„м·Ён•ң нӣ„ н—ӨнҢҢлҰ°мңјлЎң мІҳлҰ¬н•ң л©ёк· нҠңлёҢм—җ лӘЁм•„ мӢӨн—ҳм—җ мқҙмҡ©н•ҳмҳҖлӢӨ. мұ„м·Ён•ң нҳҲм•ЎмқҖ 13,000 rpm (4В°C)м—җм„ң 10분간 мӣҗмӢ¬л¶„лҰ¬ нӣ„ мғҒмёөм•Ўмқ„ мғҲ нҠңлёҢм—җ мҳ®кІЁ -80 В°Cм—җ ліҙкҙҖн•ҳмҳҖлӢӨ. к°„кіј ліөк°• лӮҙ м§Җл°©мқҖ м Ғм¶ңн•ҳм—¬ к°Ғ мЎ°м§Ғмқҳ л¬ҙкІҢлҘј мёЎм •н•ҳкі нҠңлёҢм—җ лӢҙм•„ м•ЎмІҙм§ҲмҶҢм—җ кёүлғү мӢңнӮЁ нӣ„ -80 В°Cм—җ ліҙкҙҖн•ҳмҳҖлӢӨ.
5) мғқнҷ”н•ҷм Ғ кІҖмӮ¬
нҳҲмӨ‘ нҠёлҰ¬кёҖлҰ¬м„ёлҰ¬л“ң (ETGA-200, BioAssaySystems, Hayward, CA, USA), мҙқ мҪңл ҲмҠӨн…ҢлЎӨ (K623-100, Biovision, Milpitas, CA, USA)лҠ” colorimetric assayлЎң мёЎм •н•ҳмҳҖкі , мҪ”лҘҙнӢ°мҪ”мҠӨн…ҢлЎ (corticosterone) (CSB-E0769m, Cusabio, Wuhan, China), Leptin (K0331250P, KomaBio, Seoul, Korea) л°Ҹ мқёмҠҗлҰ°(EZRMI-13K, Millipore, Billerica MA, USA)мқҖ нҡЁмҶҢл©ҙм—ӯмёЎм •лІ•(Enzyme-Linked ImmunoSorbent Assay, ELISA)мқ„ мқҙмҡ©н•ҳм—¬ microplate reader (VersaMax ELISA Microplate Reader, Molecular Devices, Sunnyvale, CA, USA)лЎң 490 nmм—җм„ң ODк°’мқ„ кө¬н•ң нӣ„ м„ӨлӘ…м„ңм—җ лӮҳмҳӨлҠ” л°©лІ•м—җ к·јкұ°н•ҳм—¬ мёЎм •н•ҳмҳҖлӢӨ.
6) кІҪкө¬лӢ№л¶Җн•ҳкІҖмӮ¬(Oral Glucose Tolerance Test, OGTT)
16мЈј мӮ¬мңЎмқҙ лҒқлӮң нӣ„ мӢӨн—ҳлҸҷл¬јмқ„ 16мӢңк°„ кёҲмӢқмӢңнӮЁ нӣ„ кј¬лҰ¬л§җмҙҲнҳҲкҙҖм—җм„ң кіөліөмғҒнғңмқҳ нҳҲм•Ўмқ„ мұ„м·Ён•ҳмҳҖлӢӨ. к°Ғ кө°м—җ 2 g/kgмқҳ нҸ¬лҸ„лӢ№ мҡ©м•Ў(Sigma Aldrich, St. Louis, MO, USA)мқ„ кІҪкө¬нҲ¬м—¬(gavage)н•ң нӣ„ 30, 60, 90, 120분 кІҪкіј мӢң кј¬лҰ¬м •л§ҘмңјлЎңл¶Җн„° мұ„нҳҲн•ҳм—¬ нҳҲлӢ№мқҳ лҶҚлҸ„ ліҖнҷ”лҘј мёЎм •н•ҳмҳҖлӢӨ.
7) м§Җл°©мЎ°м§Ғкіј к°„м—җм„ңмқҳ RT-PCR 분м„қ
к°Ғ 100mgмқҳ м§Җл°©мЎ°м§Ғкіј к°„мңјлЎңл¶Җн„° RNAлҠ” RNeasy Mini Kit (Qiagen, Hilden, Germany)лҘј мқҙмҡ©н•ҳм—¬ 분лҰ¬н•ҳмҳҖлӢӨ. Nanodrop (nanodrop 2000C, Thermo Scientific, Wilmington, DE, USA)мңјлЎң м •лҹү нӣ„ 1 Ојgмқҳ total RNAлҘј RNAлЎң Maxime RT premix kit (Intron, Seoul, Korea)лҘј мӮ¬мҡ©н•ҳм—¬ cDNAлҘј н•©м„ұн•ҳмҳҖлӢӨ. н•©м„ұн•ң cDNA (45 ng)лҠ” primer, SYBR premix Ex Taq (TaKaRa Bio Inc., Shiga, Japan)кіј нҳјн•©н•ҳм—¬ л°ҳмқ‘мӢңмј°лӢӨ. л°ҳмқ‘кіјм •мқҖ ліҖм„ұл°ҳмқ‘(95В°C) 10мҙҲ, кІ°н•©л°ҳмқ‘(50-60В°C) 5мҙҲ, м—°мһҘл°ҳмқ‘(72В°C)мҙҲлЎң 40нҡҢ л°ҳліөн•ҳмҳҖлӢӨ. лӘЁл“ л°ҳмқ‘мқҖ 3мӨ‘л°ҳліөмӢӨн—ҳмқ„ н•ҳмҳҖмңјл©° лҢҖмЎ°мң м „мһҗлЎң glyceraldehyde-3-phosphate (GAPDH)лҘј мӮ¬мҡ©н•ҳмҳҖлӢӨ. mRNA лҚ°мқҙн„°лҠ” к°Ғк°Ғмқҳ мӮ°м¶ңлҗң threshold cycle (Ct)к°’ мӨ‘ GAPDHм—җм„ң к°Ғк°Ғмқҳ мёЎм •н•ң мң м „мһҗмқҳ Ctк°’мқ„ ләҖ м°Ёмқҙк°’(О”Ct)м—җм„ң C57BL/6Jмқҳ л°ңнҳ„к°’мқ„ кё°мӨҖмңјлЎң н•ҳм—¬ О”О”Ct к°’мқ„ кө¬н•ң нӣ„ 2-(О”О”Ct) мқҳ мҲҳм№ҳлЎң л°ңнҳ„лҹүмқ„ кі„мӮ°н•ҳмҳҖлӢӨ. к°Ғ мң м „мһҗмқҳ primer м„ңм—ҙмқҖ NCBI nucleotide sequence databaseмқҳ мң м „мһҗ м„ңм—ҙ м •ліҙлҘј нҶөн•ҙ primerлҘј л””мһҗмқё н•ҳмҳҖлӢӨ. Primerл“Өмқҳ мң м „мһҗ м„ңм—ҙмқҖ Table 1кіј к°ҷлӢӨ.
TableВ 1
Primers list
8) нҶө계분м„қ
мӢӨн—ҳкІ°кіјлҠ” нҸүк· кіј н‘ңмӨҖнҺём°ЁлЎң лӮҳнғҖлӮҙм—Ҳкі , мң мқҳм„ұ кІҖм •мқ„ мң„н•ҙм„ң Prism 5.0 (GraphPad Software, La Jolla, CA, USA)лҘј мқҙмҡ©н•ҳм—¬ one-way ANOVA (analysis of variance) 분м„қмқ„ мӢӨмӢңн•ҳкі TukeyвҖҷs multiple comparison testм—җ мқҳн•ҙ p<0.05мқҳ мҲҳмӨҖм—җм„ң мӮ¬нӣ„ кІҖм •н•ҳмҳҖлӢӨ.
м—°кө¬ кІ°кіј
1. мІҙмӨ‘мҰқк°Җ, мӢқмқҙм„ӯм·Ёлҹү, мӢқмқҙнҡЁмңЁкіј мғқнҷ”н•ҷм Ғ м§Җн‘ңмқҳ ліҖнҷ”
мӢӨн—ҳкё°к°„ лҸҷм•Ҳмқҳ мІҙмӨ‘ліҖнҷ”, мӢқмқҙм„ӯм·Ёлҹүкіј мӢқмқҙнҡЁмңЁк°’мқҖ Table 2мҷҖ к°ҷлӢӨ. 비л§Ң мң лҸ„ м „ 6мЈјл № мғқмҘҗмқҳ нҸүк· мІҙмӨ‘мқҖ 17.3Вұ0.8g мқҙм—ҲмңјлӮҳ 8мЈјмқҳ мӢӨн—ҳкё°к°„ лҸҷм•Ҳ 45%мҷҖ 60% кі м§Җл°©мӢқмқҙ мӢӨн—ҳкө°(45HFD, 60HFD)л“ӨмқҖ м •мғҒмӢқмқҙкө°(CDкө°)м—җ 비н•ҳм—¬ мІҙмӨ‘мқҙ мң мқҳн•ҳкІҢ мҰқк°Җн•ҳмҳҖлӢӨ. мғқмҘҗмқҳ мҙқ мӢқмқҙм„ӯм·ЁлҹүмқҖ CDкө°мқҙ к°ҖмһҘ л§Һкі 60HFDкө°мқҙ мң мқҳн•ҳкІҢ м ҒкІҢ м„ӯм·Ён•ҳмҳҖмңјлӮҳ(F=21.04, p=.001), л°ҳлҢҖлЎң мӢқмқҙнҡЁмңЁмқҖ CDкө°мқҙ к°ҖмһҘ лӮ®кі , 45HFDкө°мқҙ лӢӨмқҢмңјлЎң лҶ’м•ҳмңјл©°, 60HFDкө°мқҙ к°ҖмһҘ лҶ’мқҖ кІғмңјлЎң лӮҳнғҖлӮ¬лӢӨ(F=6.85, p=.016). мқҙлҠ” мӢқмқҙм„ӯм·Ёлҹүмқҙ м ҒлҚ”лқјлҸ„ кіјлҸ„н•ң м§Җл°©мқҳ м„ӯм·ЁмҷҖ н•Ёк»ҳ нҡЁмңЁм ҒмңјлЎң м—җл„Ҳм§ҖлҘј мҶҢлӘЁн•ҳлҠ” лҠҘл Ҙмқҙ л–Ём–ҙм ё мІҙмӨ‘мқҙ мҰқк°Җн•ҳмҳҖмқҢмқ„ мқҳлҜён•ңлӢӨ.
TableВ 2
Effect of High Fat Diet for 8 Weeks (N=18)
| Dependent variables | Group | F | p | ||
|---|---|---|---|---|---|
| CD (n=6) | 45HFD (n=6) | 60HFD (n=6) | |||
| Weight (g) | 21.7Вұ0.5 | 22.7Вұ2.3 | 25.0Вұ1.0* | 5.88 | .015 |
| Weight gain (g) | 3.5Вұ0.6 | 5.9Вұ2.8 | 7.7Вұ1.2* | 5.46 | .028 |
| Food intake (g) | 33.2Вұ0.3 | 33.0Вұ6.0 | 26.3Вұ1.3***,### | 21.04 | <.001 |
| FER | 0.112Вұ0.020 | 0.220Вұ0.104 | 0.313Вұ0.049* | 6.85 | .016 |
| Blood glucose (mg/dL) | 154.4Вұ9.0 | 155.5Вұ32.3 | 170.6Вұ5.7* | 4.11 | .041 |
| TG (mg/dL) | 118.8Вұ32.2 | 178.9Вұ34.4* | 158.3Вұ6.8 | 4.79 | .035 |
| TC (mg/dL) | 55.3Вұ4.7 | 65.5Вұ2.9 | 137.4Вұ14.6***,### | 110.7 | <.001 |
| Insulin (ng/mL) | 0.130Вұ0.028 | 0.199Вұ0.036 | 2.111Вұ0.271***,### | 217.8 | <.001 |
| Leptin (ng/mL) | 3.9Вұ1.3 | 24.2Вұ3.1*** | 83.9Вұ10.6***,### | 199.5 | <.001 |
мӢӨн—ҳ 8мЈј нӣ„ 16мӢңк°„ кёҲмӢқмӢңнӮЁ нӣ„ нҳҲмӨ‘ мғқнҷ”н•ҷм Ғ м§Җн‘ңл“Өмқ„ 분м„қн•ҳмҳҖлӢӨ. нҳҲмӨ‘ нҠёлҰ¬кёҖлҰ¬м„ёлҰ¬л“ң, мҙқ мҪңл ҲмҠӨн…ҢлЎӨ, мқёмҠҗлҰ°кіј л ҷнӢҙмқҳ мҲҳм№ҳ лӘЁл‘җ мӢқмқҙмқҳ м§Җл°© н•Ёмң лҹүмқҙ лҶ’мқ„мҲҳлЎқ мң мқҳн•ҳкІҢ мҰқк°Җн•ҳмҳҖкі нҳҲмӨ‘ нҠёлҰ¬кёҖлҰ¬м„ёлҰ¬л“ң, л ҷнӢҙмқҳ мҲҳм№ҳлҠ” м •мғҒмӢқмқҙм—җ лҢҖн•ҙ 45HFDкө°м—җм„ңлҸ„ мң мқҳн•ҳкІҢ мҰқк°Җн•ҳмҳҖлӢӨ. н•ңнҺё, мӢқмқҙм—җ мқҳн•ң нҳҲлӢ№м°ЁмқҙлҠ” 60HFDкө°м—җм„ңл§Ң мң мқҳн•ң мҰқк°ҖлҘј ліҙмҳҖлӢӨ(F=4.11, p=.041).
мҠӨнҠёл ҲмҠӨк°Җ 비л§Ңмқҳ л°ңмғқкіј 진н–үм—җ мҳҒн–Ҙмқ„ лҒјм№ҳлҠ”м§Җ м•Ңм•„ліҙкё° мң„н•ҳм—¬ CDкө°м—җ 비н•ҳм—¬ нҒ° ліҖнҷ”к°Җ мқјм–ҙлӮҳм§Җ м•ҠмқҖ 45HFDкө°м—җкІҢ л¶ҖлҸҷ мҠӨнҠёл ҲмҠӨлҘј м Ғмҡ©(45HFD+St)н•ң нӣ„, 비л§Ңмқҳ 진н–үм—җ мҳҒн–Ҙмқ„ лҜём№ҳлҠ”м§Җ мЎ°мӮ¬н•ҳмҳҖлӢӨ. л§Өмқј 2мӢңк°„м”© 16мЈјк°„ л¶ҖлҸҷмҠӨнҠёл ҲмҠӨлҘј м Ғмҡ©н•ң кІ°кіјлҠ” лӢӨмқҢкіј к°ҷлӢӨ(Table 3). 16мЈјк°„ 45%мқҳ кі м§Җл°©мӢқмқҙлҘј кі„мҶҚ нҲ¬м—¬н•ң кІ°кіј CDкө°м—җ 비н•ҳм—¬ мІҙмӨ‘, мӢқмқҙм„ӯм·Ёлҹүкіј мғқнҷ”н•ҷм Ғ м§Җн‘ңл“Өмқҙ м§ҖмҶҚм ҒмңјлЎң мҰқк°Җн•ҳмҳҖлӢӨ. к·ёлҹ¬лӮҳ л¶ҖлҸҷ мҠӨнҠёл ҲмҠӨлҠ” мІҙмӨ‘мҰқк°Җм—җ нҒ° м°ЁмқҙлҘј ліҙмқҙм§Җ м•Ҡм•ҳлӢӨ. мӢқмқҙм„ӯм·ЁлҹүмқҖ 45HFDкө°м—җ 비н•ҙ мң мқҳн•ҳкІҢ к°җмҶҢн•ҳмҳҖмңјлӮҳ мҳӨнһҲл Ө мӢқмқҙнҡЁмңЁмқҖ мҰқк°Җн•ҳлҠ” м–‘мғҒмқ„ ліҙмқҙлӮҳ нҶөкі„м ҒмңјлЎң мң мқҳн•ҳм§ҖлҠ” м•Ҡм•ҳлӢӨ. нҳҲлӢ№, нҳҲмӨ‘ нҠёлҰ¬кёҖлҰ¬м„ёлҰ¬л“ң, мҙқ мҪңл ҲмҠӨн…ҢлЎӨ, мқёмҠҗлҰ°кіј л ҷнӢҙмқҖ 45HFDкө°м—җм„ң мң мқҳн•ҳкІҢ мҰқк°Җн•ҳмҳҖмңјлӮҳ 45HFD+Stкө°кіјлҠ” м°Ёмқҙк°Җ м—Ҷм—ҲлӢӨ. мң„мқҳ кІ°кіјл“Өмқ„ мў…н•©н• л•Ң, мҠӨнҠёл ҲмҠӨмқҳ м Ғмҡ©мқҙ л ҷнӢҙмқҳ мҰқк°ҖмҷҖ мӢқмқҙм„ӯм·Ёлҹүмқ„ к°җмҶҢмӢңнӮӨм§Җл§Ң мғҒлҢҖм ҒмңјлЎң мӢқмқҙнҡЁмңЁмқ„ мҰқк°ҖмӢңнӮҙмңјлЎңмҚЁ м—җл„Ҳм§Җ лҢҖмӮ¬к°Җ мӣҗнҷңн•ҳм§Җ м•ҠмқҢмқ„ лӮҳнғҖлӮҙкі мһҲлӢӨ. лҳҗн•ң м§Җл°©мқҳ н•Ёмң лҹүмқҙ лӮ®лҚ”лқјлҸ„ м§ҖмҶҚм Ғмқё м„ӯм·ЁлҠ” кі нҳҲлӢ№, кі м§ҖнҳҲмҰқ, кі мқёмҠҗлҰ°нҳҲмҰқ л“ұ мӮ¬лһҢмқҳ лҢҖмӮ¬мҰқнӣ„кө°м—җм„ң лӮҳнғҖлӮҳлҠ” мҰқмғҒмқҙ мқјм–ҙлӮ¬мқҢмқ„ ліҙм—¬мЈјкі мһҲлӢӨ. нҳҲмӨ‘ мҪ”лҘҙнӢ°мҪ”мҠӨн…ҢлЎ мқҳ мҲҳм№ҳлҠ” м„ё кө° лӘЁл‘җ мң мқҳн•ң ліҖнҷ”к°Җ м—Ҷм—ҲлӢӨ.
TableВ 3
Effect of High Fat Diet and Immobilization Stress for 16 Weeks (N=18)
| Dependent variables | Group | F | p | ||
|---|---|---|---|---|---|
| CD (n=6) | 45HFD (n=6) | 45HFD+St (n=6) | |||
| Weight (g) | 24.0Вұ1.5 | 29.4Вұ3.9 | 29.4Вұ4.3 | 2.42 | .131 |
| Weight gain (g) | 5.8Вұ1.8 | 12.6Вұ4.5 | 12.4Вұ5.0 | 3.69 | .068 |
| Food intake (g) | 66.4Вұ0.6 | 57.4Вұ11.6* | 50.6Вұ11.0*** | 11.91 | <.001 |
| FER | 0.097Вұ0.031 | 0.266Вұ0.100 | 0.303Вұ0.127 | 3.07 | .096 |
| Blood glucose (mg/dL) | 135.3Вұ9.0 | 162.5Вұ9.9* | 149.2Вұ16.0 | 5.04 | .028 |
| TG (mg/dL) | 117.5Вұ5.2 | 159.9Вұ23.9 | 155.7Вұ29.0 | 3.33 | .071 |
| TC (mg/dL) | 65.1Вұ5.1 | 81.4Вұ8.3* | 86.7Вұ6.1** | 8.41 | .007 |
| Insulin (ng/mL) | 0.179Вұ0.057 | 0.285Вұ0.037 | 0.211Вұ0.045 | 1.80 | .207 |
| Leptin (ng/mL) | 4.6Вұ4.2 | 40.9Вұ5.2*** | 43.4Вұ3.8*** | 100.5 | <.001 |
| Corticosterone (ng/mL) | 66.9Вұ6.0 | 60.7Вұ12.8 | 66.5Вұ7.2 | 0.51 | .617 |
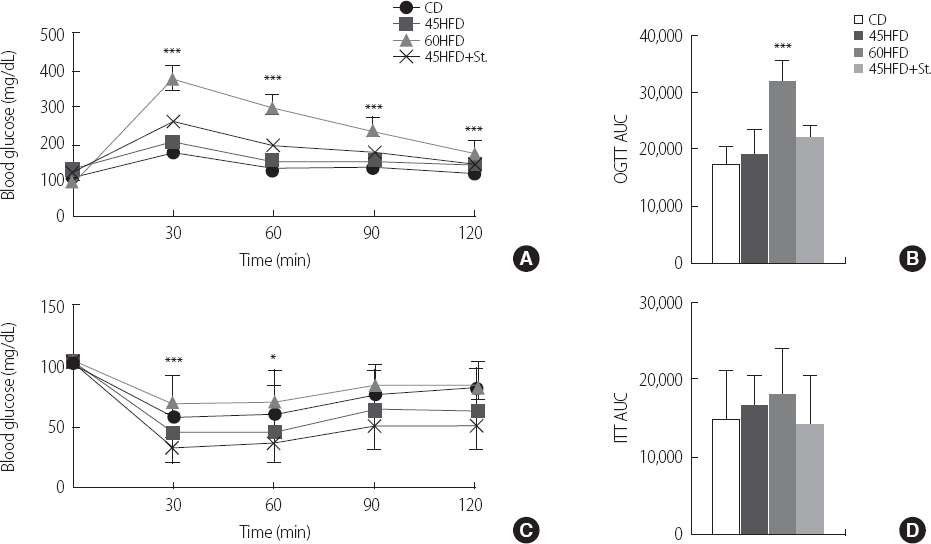
кі м§Җл°©мӢқмқҙмҷҖ мҠӨнҠёл ҲмҠӨк°Җ лӢ№л¶ҲлӮҙм„ұм—җ лҜём№ҳлҠ” мҳҒн–Ҙмқ„ мёЎм •н•ҳкё° мң„н•ҙ кІҪкө¬лӢ№л¶Җн•ҳкІҖмӮ¬(OGTT)лҘј мӢӨмӢңн•ҳмҳҖлӢӨ (Fig. 1A). кІҪкө¬лӢ№л¶Җн•ҳкІҖмӮ¬ кІ°кіј, 60HFDкө°мқҳ нҳҲмӨ‘нҸ¬лҸ„лӢ№ лҶҚлҸ„к°Җ CDкө°, 45HFDкө°, к·ёлҰ¬кі 45HFD+Stкө°м—җ 비н•ҙ кҙҖм°°н•ң лӘЁл“ мӢңк°„лҢҖм—җм„ң лҶ’мқ„ лҝҗл§Ң м•„лӢҲлқј(Fig. 1A), 120분간мқҳ мҙқнҸ¬лҸ„лӢ№лҶҚлҸ„ліҖнҷ”лҹү(Area under the curve, AUC)лҸ„ мң мқҳн•ҳкІҢ лҶ’мқҖ кІғмңјлЎң лӮҳнғҖлӮ¬лӢӨ(Fig. 1B). лҳҗн•ң мқёмҠҗлҰ° нҲ¬м—¬(0.75 U/kg) нӣ„ 2мӢңк°„ лҸҷм•Ҳмқҳ нҳҲмӨ‘нҸ¬лҸ„лӢ№ лҶҚлҸ„лҘј мёЎм •н•ҳлҠ” мқёмҠҗлҰ° лӮҙм„ұ мӢӨн—ҳ(ITT)мқ„ мӢңн–үн•ҳмҳҖмңјлӮҳ 60HFDкө°м—җм„ңл§Ң мҙҲкё° 30-60분м—җм„ң мқёмҠҗлҰ° лӮҙм„ұмқ„ ліҙм—¬ лӢ№л¶ҲлӮҙм„ұмқҳ кІ°кіјм—җ лҢҖн•ң мқјкҙҖм„ұмқ„ ліҙмҳҖмңјлӮҳ лӢӨлҘё кө°м—җм„ңлҠ” мң мқҳн•ң м°ЁмқҙлҘј ліҙмқҙм§Җ м•Ҡм•ҳлӢӨ(кІ°кіј мғқлһө).
Fig.В 1
Glucose tolerance test (GTT) and insulin tolerance test (ITT) in high fat diet-induced mice. (A) Results of the oral glucose tolerance test (OGTT) at 16 week of age showed elevated blood glucose levels in 60% high fat diet-induced mice (60HFD). (B) Area under the curve (AUC) of OGTT. (C) Results of the insulin tolerance test showed attenuated insulin responses in 60% high fat diet-induced mice. Chow diet (CD), 45% high fat diet (45HFD) or 45% high fat diet with stress (45HFD+St) shows no differences among groups. (D) Area under the curve (AUC) of ITT.
*p<.05; **p<.01; ***p<.001 compared with CD; CD=chow diet; 45HFD=45% high fat diet; 60HFD=60% high fat diet; 45HFD+St=45% high fat diet with stress.
2. мӢқмқҙм—җ л”°лҘё м§Җл°©мЎ°м§Ғ лӮҙ мң м „мһҗмқҳ ліҖнҷ”
кі м§Җл°©мӢқмқҙм—җ мқҳн•ң м§Җл°©лҢҖмӮ¬лҘј м•Ңм•„ліҙкё° мң„н•ҳм—¬ к°Ғ кө°мқҳ лӮҙмһҘм§Җл°©кіј к°„мқ„ м Ғм¶ңн•ң л’Ө кҙҖл Ё мң м „мһҗл“Өмқҳ mRNA л°ңнҳ„ м •лҸ„лҘј RT-PCRмқ„ мқҙмҡ©н•ҳм—¬ мЎ°мӮ¬н•ҳмҳҖлӢӨ(Fig. 2). м§Җл°©лҢҖмӮ¬лҠ” нҒ¬кІҢ м§Җл°©мқҳ н•©м„ұ(lipogenesis)кіј м§Җл°©мқҳ 분н•ҙ(lipolysis)лЎң кө¬л¶„н•ңлӢӨ. лӘЁл“ кө°мқҳ м§Җл°©мЎ°м§Ғм—җм„ң м§Җл°©н•©м„ұнҡЁмҶҢмқё acetyl-CoA carboxylase (Acc)мҷҖ fatty acid synthase (Fas)мқҳ mRNA л°ңнҳ„мқҖ мң мқҳн•ң м°Ёмқҙк°Җ м—Ҷм—ҲлӢӨ. к·ёлҹ¬лӮҳ м§Җ방분н•ҙ нҡЁмҶҢмқё hormone sensitive lipase (Lipe)мҷҖ adipose triglyceride lipase (Atgl)мқҳ mRNA л°ңнҳ„мқҖ 60HFDкө°м—җм„ң мң мқҳн•ҳкІҢ мҰқк°Җн•ҳлҜҖлЎң(Fig. 2A) мқҙлҠ” кі м§Җл°©мӢқмқҙм„ӯм·Ём—җ л”°лқј м§Җ방분н•ҙнҡЁмҶҢмқҳ мғқм„ұ лҳҗн•ң мҰқк°ҖмӢңнӮӨлҠ” кІғмқ„ мӢңмӮ¬н•ңлӢӨ. н•ңнҺё, мҪ”лҘҙнӢ°мҪ”мҠӨн…ҢлЎ мқҳ мҲҳмҡ©мІҙмқё nuclear receptor subfamily 3, group C, member 1 (Nr3c1) кіј к·ёмқҳ нҷңм„ұ нҡЁмҶҢмқё 11 beta-hydroxysteroid dehydrogenase type 1 (Hsd11b1)мқҳ л°ңнҳ„мқҙ 60HFDкө°м—җм„ң мң мқҳн•ҳкІҢ мҰқк°Җн•ҳмҳҖлӢӨ. Table 2мқҳ нҳҲмӨ‘ л ҷнӢҙмқҳ мҲҳм№ҳк°Җ мҰқк°Җн•ҳлҠ” кІғкіј л§Ҳм°¬к°Җм§ҖлЎң м§Җл°©м„ёнҸ¬ лӮҙ л ҷнӢҙмқҳ mRNA мҲҳм№ҳ м—ӯмӢң м§Җл°©н•Ёмң мңЁмқҙ лҶ’мқ„мҲҳлЎқ мҰқк°Җн•ҳмҳҖлӢӨ(Fig. 2B).
Fig.В 2
High fat diet alters mRNA expressions of metabolic genes in fat tissue and liver. Mice were fed with chow or high fat diet (45% or 60%) for 8 weeks. mRNA expression was determined by real-rime PCR in fat tissue (A-C) and in liver (D-I). Values were expressed as the meansВұSD standard deviation (SD) after normalization to Gapdh.
*p<.05, **p<.01 and ***p<.001 versus CD, #p<.05, ##p<.01 and ###p<.001 versus 45HFD; CD=chow diet; 45HFD=45% high fat diet; 60HFD=60% high fat diet.
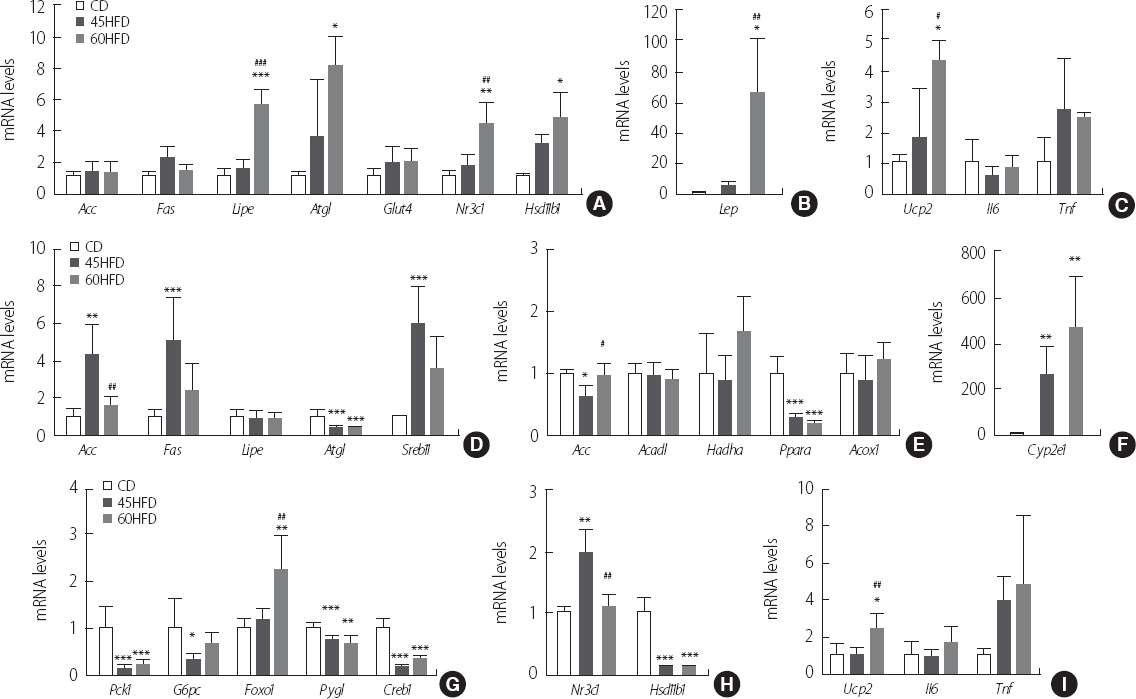
м§Җл°©мқҳ 축м ҒмқҖ мЎ°м§Ғ лӮҙ м—јмҰқл°ҳмқ‘мқ„ мҰқк°ҖмӢңнӮӨлҠ” кІғмңјлЎң м•Ңл Өм ё мһҲлӢӨ[12]. м§Җл°©мЎ°м§Ғ лӮҙ мЈјмҡ” м—јмҰқм„ұ л¬јм§Ҳмқҳ л°ңнҳ„м—¬л¶ҖлҘј мёЎм •н•ң кІ°кіј, tumor Necrosis Factor (Tnf)к°Җ кі м§Җл°©мӢқмқҙкө°л“Өм—җм„ң мҰқк°Җн•ҳлҠ” м–‘мғҒмқ„ ліҙмҳҖмңјлӮҳ мң мқҳн•ҳм§ҖлҠ” м•Ҡм•ҳмңјл©° м—јмҰқл°ҳмқ‘кіј м§Җм§Ҳ мӮ°нҷ”мҷҖ м—°кҙҖм„ұмқҙ мһҲлҠ” uncoupled protein-2 (Ucp2)лҠ” кі м§Җл°©мӢқмқҙмқҳ м§Җл°©н•Ёмң лҹүмқҙ лҶ’мқ„мҲҳлЎқ мң мқҳн•ҳкІҢ мҰқк°Җн•ҳмҳҖлӢӨ(Fig. 2C).
3. мӢқмқҙм—җ л”°лҘё к°„мЎ°м§Ғ лӮҙ мң м „мһҗмқҳ ліҖнҷ”
к°„мЎ°м§Ғ лӮҙ м§Җл°©лҢҖмӮ¬мқҳ мң м „мһҗ л°ңнҳ„мқҖ м§Җл°©мЎ°м§ҒкіјлҠ” лӢӨлҘё м–‘мғҒмңјлЎң лӮҳнғҖлӮ¬лӢӨ. 45HFDкө° к°„мЎ°м§Ғмқҳ AccмҷҖ Fas, к·ёлҰ¬кі мқҙл“Ө мң м „мһҗмқҳ м „мӮ¬мқёмһҗмқё sterol regulatory element transcription factor (Srebf1)мқҳ л°ңнҳ„мқҙ мҰқк°Җн•ҳлҠ” л°ҳл©ҙ, Atglкіј к°ҷмқҙ м§Җл°© 분н•ҙм—җ кҙҖл Ёлҗң мң м „мһҗмқҳ л°ңнҳ„мқҖ мң мқҳн•ҳкІҢ к°җмҶҢн•ҳмҳҖлҠ”лҚ°(Fig. 2D), мқҙлҠ” кі м§Җл°©мӢқмқҙм—җ л”°лқј к°„м—җ м§Җл°©мқ„ н•©м„ұн•ҳкі м¶•м Ғн•ҳлҠ” кіјм •мңјлЎң ліҙмқёлӢӨ. м§Җл°©мқҖ лІ нғҖ мӮ°нҷ”кіјм •(ОІ-oxidation)мқ„ нҶөн•ҳм—¬ м§Җл°©мқ„ 분н•ҙн•ҳм—¬ м—җл„Ҳм§ҖмӣҗмңјлЎң м „нҷҳн•ҳлҠ”лҚ° мқҙл•Ң, м§Җл°©мӮ°мқ„ лҜёнҶ мҪҳл“ңлҰ¬м•„лЎң м „лӢ¬н•ҳм—¬ мЈјлҠ” carnitine palmitoyltransferase 1a (Cpt1a)мқҳ л°ңнҳ„мқҙ л‘җ кі м§Җл°©мӢқмқҙкө°м—җм„ң к°җмҶҢн•ҳкі , long-chain L-3-hydroxyacyl-coenzyme A dehydrogenase (HADHОұ), acyl-Coenzyme A dehydrogenase, long-chain (Acadl), peroxisomal acyl-coenzyme A oxidase 1 (Acox1)кіј к°ҷмқҙ м§Җл°©мӮ°мқҳ мӮ°нҷ”кіјм •м—җ кҙҖм—¬н•ҳлҠ” мң м „мһҗмқҳ л°ңнҳ„м—җлҠ” ліҖнҷ”к°Җ м—Ҷм—ҲлӢӨ. нҠ№нһҲ, peroxisome proliferators-activated receptor (Ppara)мқҳ mRNA л°ңнҳ„мқҙ кі м§Җл°©мӢқмқҙкө°мқҳ к°„мЎ°м§Ғм—җм„ң нҳ„м Җн•ҳкІҢ л–Ём–ҙм§ҖлҠ” кІғмңјлЎң ліҙм•„ мқҙлҠ” м§Җл°©мӮ°мқҳ лҜёнҶ мҪҳл“ңлҰ¬м•„ лӮҙ мң мһ…кіј м§Җл°©мӮ° мӮ°нҷ”м—җ мһҘм• к°Җ мқјм–ҙлӮҳлҠ” кІғмқ„ мқҳлҜён•ҳл©° мқҙлҠ” Pparaмқҳ м§Ғм ‘м Ғмқё нғҖкІҹмң м „мһҗмқё Cpt1a л°ңнҳ„мқҳ к°җмҶҢмҷҖлҸ„ мқјкҙҖм„ұмқ„ ліҙмқҙкі мһҲлӢӨ(Fig. 2E). м§Җл°©мӮ°мқҳ лІ нғҖ мӮ°нҷ” кҙҖл Ё мң м „мһҗл“Өмқҳ л°ңнҳ„м—җ нҒ° ліҖнҷ”к°Җ мқјм–ҙлӮҳм§Җ м•ҠлҠ” л°ҳл©ҙ, лҜём„ёмҶҢмІҙ(microsome)м—җ мқҳн•ң мҳӨл©”к°Җ мӮ°нҷ”(ОІ-oxidation)м—җ кҙҖм—¬н•ҳлҠ” cytochrome P450 2E1 (Cyp2e1)мқҳ л°ңнҳ„мқҖ кі м§Җл°©мӢқмқҙмқҳ м§Җл°©н•Ёмң лҹүмқҙ лҶ’мқ„мҲҳлЎқ мҰқк°Җн•ҳмҳҖлӢӨ(Fig. 2F). Fig. 2D-Fмқҳ кІ°кіјлҘј мў…н•©н•ҳм—¬ ліј л•Ң кі м§Җл°©мӢқмқҙлҠ” к°„мЎ°м§ҒлӮҙ м§Җл°©мқҳ н•©м„ұмқ„ мҰқк°Җн•ҳм—¬ 축м Ғн•ҳм§Җл§Ң мқҙлҘј м—җл„Ҳм§ҖмӣҗмңјлЎң м „нҷҳн•ҳм—¬ мҶҢ비мӢңнӮӨлҠ” лҜёнҶ мҪҳл“ңлҰ¬м•„ лӮҙ лІ нғҖ мӮ°нҷ”лҘј нҷңм„ұнҷ”мӢңнӮӨм§Җ лӘ»н•ҳкі кіјлҸ„н•ң м§Җл°©мӮ°мқҳ 축м ҒмңјлЎң мқјм–ҙлӮҳлҠ” лҢҖмІҙмӮ°нҷ”мһ‘мҡ©мқё лҜём„ёмҶҢмІҙ лӮҙ мҳӨл©”к°Җ мӮ°нҷ”к°Җ мқјм–ҙлӮҳлҠ” кІғмқ„ мӢңмӮ¬н•ңлӢӨ.
к°„мЎ°м§Ғ лӮҙ лӢ№лҢҖмӮ¬мқҳ ліҖнҷ”лҘј мӮҙнҺҙліҙкё° мң„н•ҳм—¬ лӢ№мӢ мғқкіјм • (glyconeogenesis)м—җ кҙҖм—¬н•ҳлҠ” мң м „мһҗмқҳ л°ңнҳ„мқ„ нҷ•мқён•ҳмҳҖлӢӨ. phosphoenolpyruvate carboxykinase 1 (Pck1)кіј glucose-6-phosphatase (G6pc)мқҳ л°ңнҳ„мқҖ кі м§Җл°©мӢқмқҙкө°м—җм„ң мң мқҳн•ҳкІҢ к°җмҶҢн•ҳмҳҖкі , м „мӮ¬мқёмһҗлЎң мһ‘мҡ©н•ҳлҠ” cAMP responsive element binding protein 1 (Creb1) лҳҗн•ң кі м§Җл°©мӢқмқҙкө°м—җм„ң к°җмҶҢн•ҳмҳҖлӢӨ. к·ёлҹ¬лӮҳ лҳҗ лӢӨлҘё м „мӮ¬мқёмһҗмқё forkhead box protein O1 (Foxo1)мқҖ 60HFDкө°м—җм„ң мҰқк°Җн•ҳмҳҖлӢӨ. к°„м—җм„ң кёҖлҰ¬мҪ”кІҗмқ„ мқҙмҡ©н•ҳм—¬ лӢ№мӣҗ분н•ҙкіјм •(glycogenolysis)м—җ мһ‘мҡ©н•ҳлҠ” мң м „мһҗ liver glycogen phosphorylase (Pygl)мқҳ л°ңнҳ„ м—ӯмӢң кі м§Җл°©мӢқмқҙм—җм„ң мң мқҳн•ҳкІҢ к°җмҶҢн•ҳмҳҖлӢӨ(Fig. 2G).
н•ңнҺё, к°„м—җм„ңмқҳ Nr3c1кіј Hsd11b1мқҖ м§Җл°©мЎ°м§ҒкіјлҠ” лӢӨлҘё м–‘мғҒмқ„ ліҙмҳҖлӢӨ. Nr3c1мқҖ 45HFDкө°м—җм„ң мң мқҳн•ң мҰқк°ҖлҘј ліҙмқҙлӢӨ 60HFDкө°м—җм„ңлҠ” к°җмҶҢлҘј н•ҳмҳҖкі , Hsd11b1мқҖ лӘЁл“ кі м§Җл°©мӢқмқҙкө°м—җм„ң нҳ„м Җн•ҳкІҢ к°җмҶҢн•ҳмҳҖлӢӨ(Fig. 2-H). Ucp2мқҳ л°ңнҳ„мқҖ м§Җл°©мЎ°м§Ғм—җм„ңмҷҖ л§Ҳм°¬к°Җм§ҖлЎң 60HFDкө°м—җм„ң мҰқк°Җн•ҳмҳҖмңјлӮҳ Il6лӮҳ Tnfмқҳ кІҪмҡ° мҰқк°Җн•ҳлҠ” кІҪн–Ҙмқҙ лӮҳнғҖлӮҳм§Җл§Ң мң мқҳн•ң м°ЁмқҙлҘј ліҙмқҙм§Җ м•Ҡм•ҳлӢӨ(Fig. 2I). мң„мқҳ кІ°кіјлҘј мў…н•©н•ҳм—¬ ліј л•Ң кі м§Җл°©мӢқмқҙм—җ мқҳн•ң к°„мЎ°м§Ғ лӮҙ м§Җл°©мқҳ н•©м„ұмқҖ м Ғм Ҳн•ң лҜёнҶ мҪҳл“ңлҰ¬м•„мқҳ лІ нғҖ мӮ°нҷ”лҘј м Җн•ҙн•ҳкі лҜём„ёмҶҢмІҙмқҳ мҳӨл©”к°Җ мӮ°нҷ”лҘј мҰқк°ҖмӢңнӮӨлҠ” кІ°кіјлҘј к°Җм ёмҳӨл©° нҳҲмӨ‘ нҸ¬лҸ„лӢ№ мҲҳм№ҳк°Җ мҰқк°Җн•Ём—җ л”°лқј лӢ№мӢ мғқкіјм •мқҙлӮҳ лӢ№мӣҗ분н•ҙкіјм •мқҖ м–өм ңлҗҳлҠ” кІғмқ„ м•Ң мҲҳ мһҲлӢӨ.
4. л¶ҖлҸҷ мҠӨнҠёл ҲмҠӨ(immobilization stress)м—җ мқҳн•ң мң м „мһҗ л°ңнҳ„мқҳ ліҖнҷ”
45% кі м§Җл°©мӢқмқҙмҷҖ н•Ёк»ҳ л¶ҖлҸҷ мҠӨнҠёл ҲмҠӨм—җ 16мЈј лҸҷм•Ҳ л…ём¶ң мӢңмј°мқ„ л•Ң мң м „мһҗмқҳ л°ңнҳ„ ліҖнҷ”лҘј 비көҗ, нҷ•мқён•ҳмҳҖлӢӨ(Fig. 3). м§Җл°©мЎ°м§Ғм—җм„ң м§Җл°©н•©м„ұм—җ кҙҖм—¬н•ҳлҠ” мң м „мһҗ AccмҷҖ Fasмқҳ л°ңнҳ„мқҖ 45HFDкө°кіј 45HDS+Stкө° лӘЁл‘җ мң мқҳн•ҳкІҢ к°җмҶҢн•ҳмҳҖкі , м§Җ방분н•ҙмҷҖ кҙҖл Ёлҗң мң м „мһҗл“ӨмқҙлӮҳ Glut4мқҳ л°ңнҳ„м—җлҠ” м°Ёмқҙк°Җ м—Ҷм—ҲлӢӨ. 45HFDкө°м—җм„ң мҰқк°Җн•ң Nr3c1кіј Hsd11b1мқҳ л°ңнҳ„мқҖ мҠӨнҠёл ҲмҠӨм Ғмҡ©м—җ мқҳн•ҙ мң мқҳн•ң м°ЁмқҙлҘј ліҙмқҙм§Җ м•ҠмңјлӮҳ м җм°Ё л°ңнҳ„мқҙ мҰқк°Җн•ҳлҠ” м–‘мғҒмқ„ ліҙмҳҖлӢӨ(Fig. 3A). л ҷнӢҙмқҳ мң м „мһҗ л°ңнҳ„мқҖ CDкө° лҢҖ비 45HFDкө°м—җм„ң мң мқҳн•ҳкІҢ мҰқк°Җн•ҳмҳҖмңјлӮҳ мҠӨнҠёл ҲмҠӨ м Ғмҡ©мқҙ нҒ° мҳҒн–Ҙмқ„ лҜём№ҳм§Җ м•ҠмқҖ кІғмңјлЎң ліҙмқҙл©°(Fig. 3B), мӮ°нҷ” мҠӨнҠёл ҲмҠӨм—җ кҙҖм—¬н•ҳлҠ” Ucp2мҷҖ м—јмҰқ н‘ңм§Җмһҗмқё Il6лӮҳ Tnfмқҳ л°ңнҳ„мқҖ мҠӨнҠёл ҲмҠӨ м Ғмҡ©мқ„ л°ӣмқҖ 45HFD+Stкө°м—җм„ң мң мқҳн•ҳкІҢ к°җмҶҢн•ҳмҳҖлҠ”лҚ°, мқҙлҠ” л¶ҖлҸҷмҠӨнҠёл ҲмҠӨм—җ мқҳн•ң Nr3c1кіј Hsd11b1 мң м „мһҗ л°ңнҳ„мқҳ мҰқк°Җк°Җ мЎ°м§Ғ лӮҙ мҪ”нӢ°мҶ” нҷңм„ұмқ„ лҶ’мқё кІғмңјлЎң ліҙмқҙл©° Il6лӮҳ Tnfмқҳ к°җмҶҢлҠ” мҪ”нӢ°мҶ” нҷңм„ұ мҰқк°Җм—җ мқҳн•ң л©ҙм—ӯм–өм ңнҡЁкіјлҘј л°ҳмҳҒн•ҳлҠ” кІғмңјлЎң ліҙмқёлӢӨ(Fig. 3C).
Fig.В 3
Immobilization stress alters mRNA expressions of metabolic genes in fat tissue and liver. Physical immobilization was used to cause chronic stress. 45% high fat diet-induced mice received immobilization stress for 2 hr daily for 16 weeks. mRNA expression was determined by real-rime PCR in fat tissue (A-C) and in liver (D-I). Values were expressed as the meansВұSD standard deviation (SD) after normalization to Gapdh.
*p<.05, **p<.01 and ***p<.001 versus CD, #p<.05, ##p<.01 and ###p<.001 versus 45HFD; CD=chow diet; 45HFD=45% high fat diet; 60HFD=60% high fat diet.
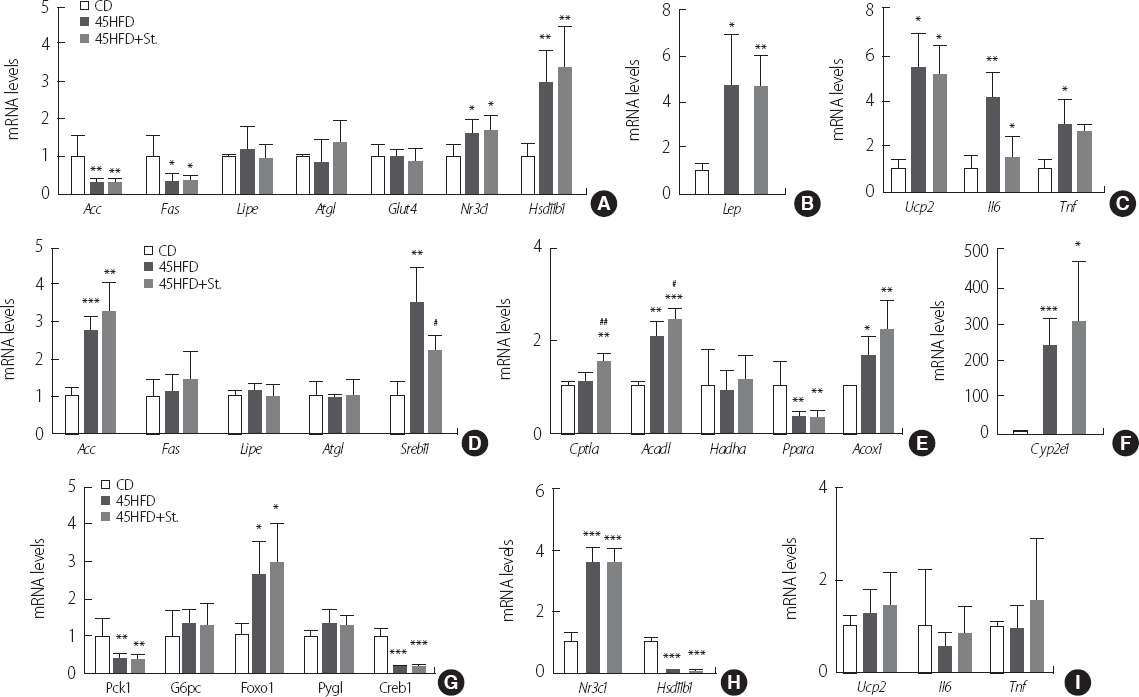
л¶ҖлҸҷ мҠӨнҠёл ҲмҠӨмқҳ м Ғмҡ©мқҖ к°„ мЎ°м§Ғ лӮҙ м§Җл°©мӮ°мқҳ мӮ°нҷ”кіјм •м—җ мҳҒн–Ҙмқ„ мЈјлҠ” кІғмңјлЎң лӮҳнғҖлӮ¬лӢӨ. 16мЈјк°„ кі м§Җл°©мӢқмқҙлҘј м„ӯм·Ён•ң мҘҗмқҳ к°„мЎ°м§Ғм—җм„ң м§Җл°©мқҳ н•©м„ұкіј 분н•ҙлҠ” Accмқҳ л°ңнҳ„мқ„ м ңмҷён•ҳкі лҠ” нҒ° ліҖнҷ”к°Җ м—Ҷмқ„ лҝҗ м•„лӢҲлқј, м§Җл°©н•©м„ұмқҳ мЈјмҡ” м „мӮ¬мқёмһҗмқё Srebf1мқҳ л°ңнҳ„мқҙ мҠӨнҠёл ҲмҠӨ м Ғмҡ©м—җ мқҳн•ҙ к°җмҶҢлҗҳм—ҲмқҢмқ„ нҷ•мқён•ҳмҳҖлӢӨ(Fig. 3D). мқҙм—җ 비н•ҳм—¬ м§Җл°©мқҳ лІ нғҖ мӮ°нҷ”мҷҖ мҳӨл©”к°Җ мӮ°нҷ”лҠ” лҚ” нҷңл°ңнһҲ мқјм–ҙлӮҳ Cpt1a, Acadl, Acox1мқҳ л°ңнҳ„мқҙ мң мқҳн•ҳкІҢ мҰқк°Җн•ҳмҳҖмңјл©°(Fig. 3E) Cyp2e1мқҳ л°ңнҳ„мқҖ мҠӨнҠёл ҲмҠӨм—җ мқҳн•ҳм—¬ лҚ” нҷңм„ұнҷ” лҗҳлҠ” кІғмқ„ м•Ң мҲҳ мһҲм—ҲлӢӨ(Fig. 2E, Fig. 2F). лӢ№лҢҖмӮ¬мҷҖ кҙҖл Ёлҗң мң м „мһҗл“ӨмқҖ лӢ№мӢ мғқкіјм •кіј лӢ№мӣҗ분н•ҙкіјм • лӘЁл‘җ 8мЈј мӢңм җкіј 비көҗн•ҳм—¬ нҒ° ліҖнҷ”к°Җ ліҙмқҙм§Җ м•Ҡм•ҳмңјл©°(Fig. 3G) мҠӨнҠёл ҲмҠӨк°Җ мқҙлҹ¬н•ң мң м „мһҗл“Өмқҳ л°ңнҳ„м—җ мҳҒн–Ҙмқ„ лҜём№ҳм§Җ м•Ҡм•ҳлӢӨ. Nr3c1кіј Hsd11b1 мқҳ л°ңнҳ„ м—ӯмӢң 8мЈј мӢңм җкіј м°Ёмқҙк°Җ лӮҳнғҖлӮҳм§Җ м•Ҡм•ҳмңјл©°(Fig. 3H), м§Җл°©мЎ°м§ҒкіјлҠ” лӢ¬лҰ¬ м—јмҰқ мӮ¬мқҙнҶ м№ҙмқё Il6мҷҖ Tnf к·ёлҰ¬кі Ucp2мқҳ л°ңнҳ„мқҖ кі м§Җл°©мӢқмқҙлӮҳ мҠӨнҠёл ҲмҠӨм—җ мқҳн•ң ліҖнҷ”к°Җ м—Ҷм—ҲлӢӨ(Fig. 3I).
5. мӢқмқҙм—җ л”°лҘё мғқмІҙмӢңкі„ мң м „мһҗ л°ңнҳ„мқҳ ліҖнҷ”
ліё м—°кө¬лҠ” кі м§Җл°©мӢқмқҙмҷҖ мҠӨнҠёл ҲмҠӨк°Җ м§Җл°©мЎ°м§Ғкіј к°„мЎ°м§Ғмқҳ мғқмІҙмӢңкі„ мң м „мһҗмқҳ л°ңнҳ„м—җ ліҖнҷ”лҘј мқјмңјнӮӨлҠ”м§Җ мң м „мһҗмқҳ mRNAмқҳ RT-PCRмқ„ нҶөн•ҳм—¬ нҷ•мқён•ҳмҳҖлӢӨ(Fig. 4). м§Җл°©мЎ°м§Ғм—җм„ңлҠ” Clock, aryl hydrocarbon receptor nuclear translocator-like protein (Arntl;л§Ҳмҡ°мҠӨмқҳ BMAL1 мң м „мһҗ), Cry1, Per1, Per2, Per3 лӘЁл‘җ кі м§Җл°© н•Ёмң лҹүмқҙ л§Һмқ„мҲҳлЎқ mRNAмқҳ л°ңнҳ„мқҙ мҰқк°Җн•ҳлҠ” кІҪн–Ҙмқ„ ліҙмҳҖкі нҠ№нһҲ Clock, Arntlкіј Per2мқҳ л°ңнҳ„мқҙ мң мқҳн•ҳкІҢ мҰқк°Җн•ҳмҳҖлӢӨ(Fig. 4A). к°„мЎ°м§Ғ лӮҙмқҳ мғқмІҙмӢңкі„ мң м „мһҗмқҳ л°ңнҳ„ м—ӯмӢң кі м§Җл°©мӢқмқҙм—җ л”°лқј ліҖнҷ”к°Җ мһҲм—ҲлҠ”лҚ° 45HFDкө°м—җм„ң Clock, Cry1мқҳ л°ңнҳ„мқҖ к°җмҶҢн•ҳкі Per 1-3мқҳ л°ңнҳ„мқҖ мҰқк°Җн•ҳмҳҖлӢӨ. мқҙлҠ” Clock/Arntl ліөн•©мІҙ(complex)к°Җ Cry/Perмқҳ ліөн•©мІҙмҷҖ м„ңлЎң мғҒнҳёмһ‘мҡ©н•ҳлҜҖлЎң мқҙм—җ л”°лҘё ліҖнҷ”мқё кІғмңјлЎң ліҙмқёлӢӨ. мқҙлҹ¬н•ң л°ңнҳ„мқҳ ліҖнҷ”лҠ” 60HFDкө°м—җм„ң л°ҳлҢҖ м–‘мғҒмқ„ ліҙм—¬ Clock, Arntl, Cry1мқҳ л°ңнҳ„мқҙ мҰқк°Җн•ҳлҠ” л°ҳл©ҙ, Per1-3мң м „мһҗмқҳ л°ңнҳ„мқҖ м§Җл°© н•Ёмң лҹүмқҙ лҶ’мқ„мҲҳлЎқ(60HFD) к°җмҶҢн•ҳмҳҖлӢӨ(Fig. 4B).
Fig.В 4
High fat diet and immobilization stress alters mRNA expression of circadian clock genes in fat and liver. mRNA expression of circadian clock genes in fat (A) and liver (B) at 8 weeks of age. mRNA expression of circadian clock genes in fat (A) and liver (B) at 16 weeks of age. Values were expressed as the meansВұSD standard deviation (SD) after normalization to Gapdh.
*p<.05, **p<.01 and ***p<.001 versus CD, #p<.05, ##p<.01 and ###p<.001 versus 45HFD; CD, chow diet; 45HFD, 45% high fat diet; 60HFD, 60% high fat diet; 45HFD+St, 45% high fat diet with stress.
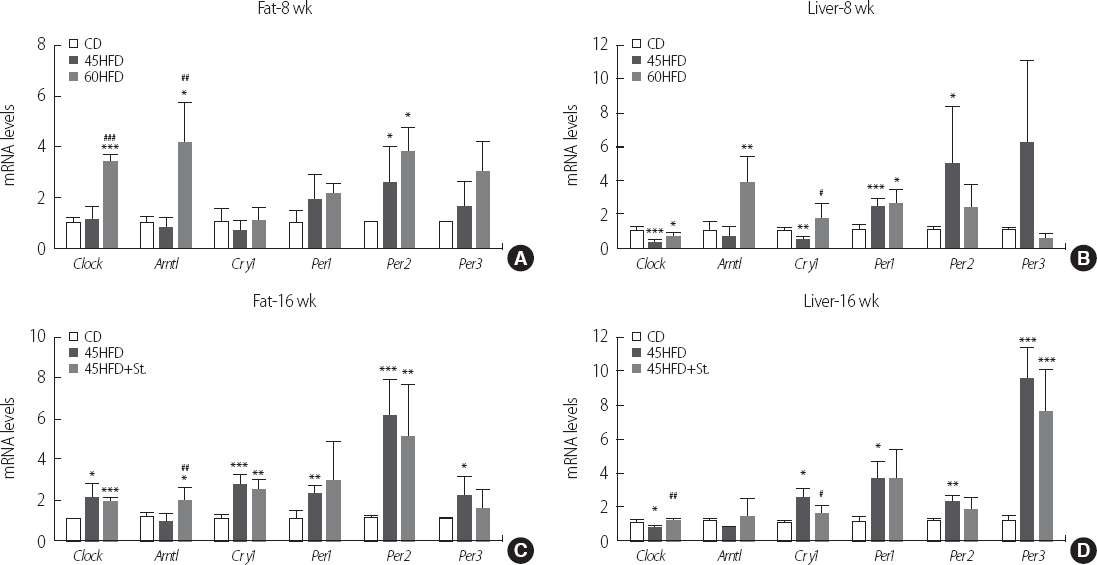
н•ңнҺё, 16мЈј лҸҷм•Ҳ 45% кі м§Җл°©мӢқмқҙлҘј м„ӯм·Ён•ҳкІҢ лҗҳл©ҙ 8мЈјмҷҖлҠ” лӢ¬лҰ¬ м§Җл°©мЎ°м§Ғмқҳ лҢҖл¶Җ분мқҳ мң м „мһҗ л°ңнҳ„мқҙ мң мқҳн•ҳкІҢ мҰқк°Җн•ҳмҳҖлӢӨ. нҠ№нһҲ, 45HFDкө°мқҳ Per1-3мқҳ л°ңнҳ„мқҙ 8мЈј мӢңкё°м—җ 비н•ҙ CDкө°кіј 비көҗн•ҳмҳҖмқ„ л•Ң м°Ёмқҙк°Җ нҒ¬лҜҖлЎң кі м§Җл°©мӢқмқҙмқҳ кё°к°„мқҙ кёёмҲҳлЎқ мң м „мһҗл“Өмқҳ л°ңнҳ„мқҙ лҚ” мҰқк°Җн•ҳлҠ” кІғмқ„ м•Ң мҲҳ мһҲлӢӨ. н•ң нҺё, мҠӨнҠёл ҲмҠӨлҘј м Ғмҡ©н•ҳмҳҖмқ„ л•Ң м§Җл°© лӮҙ ArntlмқҖ мң мқҳн•ҳкІҢ мҰқк°Җн•ң л°ҳл©ҙ, к·ё мҷёмқҳ мғқмІҙмӢңкі„ мң м „мһҗл“Өмқҳ л°ңнҳ„мқҖ мҠӨнҠёл ҲмҠӨм—җ мқҳн•ҙ к°җмҶҢн•ҳлҠ” кІғмқҙ л°ңкІ¬лҗҳм—ҲлӢӨ(Fig. 4C).
к°„мЎ°м§Ғмқҳ кІҪмҡ°, 8мЈј мӢңкё°мҷҖ мң мӮ¬н•ҳкІҢ Clock/Arntlмқҳ л°ңнҳ„мқҖ к°җмҶҢн•ҳкі , Per1-3мқҳ л°ңнҳ„мқҖ мҰқк°Җн•ҳмҳҖлӢӨ. 45HFD+Stкө°м—җм„ң Clockкіј Cry1мқҳ л°ңнҳ„м—җ мң мқҳн•ң ліҖнҷ”к°Җ мһҲм—ҲмңјлӮҳ к·ё мҷёмқҳ мң м „мһҗм—җлҠ” нҒ° мҳҒн–Ҙмқҙ лӮҳнғҖлӮҳм§Җ м•Ҡм•ҳмңјлӮҳ(Fig. 4D). 45HFD+Stкө°мқҳ Per1-3мқҳ л°ңнҳ„мқҙ 60% HFDкө°м—җм„ңмҷҖ к°ҷмқҙ 45HFDкө°м—җ 비көҗн•ҳм—¬ к°җмҶҢн•ҳлҠ” кІғмқ„ м•Ң мҲҳ мһҲм—ҲлӢӨ(Fig. 4D). н•ң к°Җм§Җ мЈјлӘ©н• м җмқҖ м§Җл°©мЎ°м§ҒмқҖ Per2к°Җ, к°„мЎ°м§ҒмқҖ Per3мқҳ л°ңнҳ„м—җ нҒ° ліҖнҷ”лҘј ліҙм—¬ мһҘкё°лӮҳ мЎ°м§Ғм—җ л”°лқј мЈјмҡ” м—ӯн• мқ„ н•ҳлҠ” мң м „мһҗк°Җ лӢӨлҘҙкІҢ мһ‘лҸҷн•ҳлҠ” кІғмңјлЎң ліҙмқёлӢӨ.
л…јмқҳ
ліё м—°кө¬лҠ” 비л§Ңмқҙ л°ңмғқн•ҳмҳҖмқ„ л•Ң мқјм–ҙлӮҳлҠ” лҢҖмӮ¬м„ұ ліҖнҷ”лҘј ліҙкё° мң„н•ҳм—¬ мӮ¬лһҢкіј к°ҖмһҘ мң мӮ¬н•ҳкІҢ лӮҳнғҖлӮҳлҠ” C57BL/6J л§Ҳмҡ°мҠӨм—җкІҢ ліҙнҺём ҒмңјлЎң мӮ¬мҡ©н•ҳлҠ” 45%мҷҖ 60% кі м§Җл°©мӢқмқҙлҘј к°Ғк°Ғ кіөкёүн•ҳм—¬ 10%мқҳ м§Җл°©мқ„ н•Ёмң н•ҳкі мһҲлҠ” м •мғҒмӢқмқҙ(CD)лҘј кіөкёүн•ҳмҳҖмқ„ л•ҢмҷҖ м–ҙл–Ө ліҖнҷ”к°Җ мқјм–ҙлӮҳлҠ”м§Җ 비көҗ, 분м„қн•ҳмҳҖлӢӨ[13]. м§Җл°©н•Ёмң лҹүмқҙ лҶ’кұ°лӮҳ кіјлҸ„н•ң м№јлЎңлҰ¬ м„ӯм·Ём—җ мқҳн•ҙ 비л§Ңмқҙ л°ңмғқн•ҳл©ҙ м§Җл°©лҢҖмӮ¬мқҳ мһҘм• к°Җ мқјм–ҙлӮҳл©ҙм„ң к°„мЎ°м§Ғм—җ кіјлҸ„н•ң м§Җл°©мқҙ 축м Ғмқҙ лҗҳлҠ” м§Җл°©к°„мҰқ лҳҗлҠ” 비м•ҢмҪңм„ұ м§Җл°©к°„м§Ҳнҷҳмқҳ л°ңмғқмқҙ мҰқк°Җн•ҳкІҢ лҗңлӢӨ. м§Җл°©к°„мқҳ мҰқк°Җ кё°м „мқҖ мІ«м§ё, мӢқмқҙ м§Җл°© нҳ№мқҖ м№јлЎңлҰ¬мқҳ кіјлӢӨ м„ӯм·Ём—җ мқҳн•ң к°„мңјлЎң м§Җл°© мң мһ…(inflow)мқҳ мҰқк°Җ, л‘ҳм§ё, м§Җл°©мЎ°м§Ғ(adipose tissue)м—җм„ң м§Җ방분н•ҙ(lipolysis)мқҳ мҰқк°ҖлЎң мқён•ң к°„мңјлЎңмқҳ кіјлҸ„н•ң мң лҰ¬м§Җл°©мӮ°(free fatty acids)мқҳ мң мһ…, м…Ӣм§ё, к°„м—җм„ң мӢ к·ң м§Җл°©н•©м„ұ(de novo lipogenesis)мқҳ мҰқк°Җ, л„·м§ё, к°„м—җм„ң мҙҲм Җл°ҖлҸ„мҪңл ҲмҠӨн…ҢлЎӨ(Very Low Density Lipoprotein)мқ„ нҶөн•ң TG мң м¶ң(outflow)мқҳ к°җмҶҢ, к·ёлҰ¬кі лӢӨм„Ҝм§ёлЎң к°„м—җм„ң м§Җл°©мӮ° мӮ°нҷ”(oxidation)мқҳ к°җмҶҢ, мҰү кіөліө мӨ‘ лӢ№мӢ мғқ(gluconeogenesis) л“ұмқҳ мЈјмҡ” м—җл„Ҳм§Җмӣҗ мӮ¬мҡ©мқҳ к°җмҶҢ л“ұмқҙлӢӨ[14]. ліё м—°кө¬лҘј нҶөн•ҙ лӮҳнғҖлӮң кІ°кіјл“ӨмқҖ м§Җл°©к°„мқҙ л°ңмғқн•ҳлҠ” кё°м „мқ„ мҲңм°Ём ҒмңјлЎң мҰқлӘ…н•ҳкі мһҲлӢӨ. 45% лҳҗлҠ” 60%мқҳ кі м§Җл°©мӢқмқҙлҘј кіөкёүн•ҳкі 8мЈјк°Җ м§ҖлӮң мӢңм җм—җ м§Җл°©мЎ°м§Ғм—җм„ңлҠ” м§Җл°©мқҳ н•©м„ұліҙлӢӨлҠ” м§Җл°© 분н•ҙм—җ кҙҖм—¬н•ҳлҠ” нҡЁмҶҢл“Ө, LipeмҷҖ Atglмқҳ л°ңнҳ„мқҙ мң мқҳн•ҳкІҢ мҰқк°Җн•ҳмҳҖлӢӨ. LipeмҷҖ AtglмқҖ м§Җл°©мЎ°м§Ғм—җм„ң нҠёлҰ¬кёҖлҰ¬м„ёлҰ¬л“ңлҘј лӢӨмқҙм•„мӢӨкёҖлҰ¬м„ёлЎӨ(diacylglycerol;DAG)кіј мң лҰ¬м§Җл°©мӮ°(free fatty acids)мңјлЎң 분н•ҙн•ҳлҠ” нҡЁмҶҢмқҙлӢӨ. лҸҷмӢңм—җ к°„м—җм„ңлҠ” м§Җл°©мғқм„ұмқҳ м „мӮ¬мҡ”мҶҢ(transcription factor)мқё Srebf1мқ„ нҶөн•ң Fas, AccмҷҖ к°ҷмқҖ м§Җм§Ҳ н•©м„ұ нҡЁмҶҢмқҳ л°ңнҳ„мқҙ мҰқк°Җн•ҳкі к·ё кІ°кіјлЎң нҳҲмӨ‘ м§Җм§Ҳ м§Җн‘ңмқҳ мҰқк°Җк°Җ мқјм–ҙлӮЁмқ„ нҷ•мқён•ҳмҳҖлӢӨ. м •мғҒм Ғмқё лҢҖмӮ¬кіјм •м—җм„ң мқёмҠҗлҰ°мқҖ м§Җл°©мЎ°м§Ғм—җм„ң м§Җл°© 분н•ҙлҘј м–өм ңн•ҳм§Җл§Ң ліё м—°кө¬м—җм„ңмҷҖ 60HFDкө°м—җм„ңлҠ” нҳҲмӨ‘ мқёмҠҗлҰ°мқҖ мҰқк°Җн•ҳлӮҳ мқёмҠҗлҰ° м Җн•ӯм„ұмқҙ мҲҳл°ҳлҗҳл©ҙм„ң м§Җл°©мЎ°м§Ғмқҳ м§Җм§Ҳ 분н•ҙк°Җ мҙү진лҗҳкі (Fig. 2A), 분н•ҙлҗң мң лҰ¬м§Җл°©мӮ°мқҖ к°„мңјлЎң мң мһ…лҗҳлҠ” лҸҷмӢңм—җ Srebf1мқҳ мң м „мһҗ л°ңнҳ„мқ„ мҰқк°ҖмӢңнӮӨлҠ” кІғмқ„ нҷ•мқён• мҲҳ мһҲм—ҲлӢӨ[15]. мқҙлҹ¬н•ң мқёмҠҗлҰ° м Җн•ӯм„ұкіј к°„м—җм„ң мӨ‘м„ұм§Җл°©мқҳ 축м Ғм—җ лҢҖн•ң Srebf1мқҳ м—ӯн• мқҖ лӢӨлҘё лҸҷл¬јлӘЁлҚёмқ„ нҶөн•ҙм„ңлҸ„ мҰқлӘ…лҗҳм—ҲлӢӨ. кі лҸ„비л§Ңкіј мқёмҠҗлҰ° м Җн•ӯм„ұмқҳ нҠ№м§•мқё ob/ob miceм—җм„ң м§Җл°©к°„ лі‘ліҖмқҙ кҙҖм°°лҗҳм—Ҳкі , ob/ob miceмқҳ Srebf1 мң м „мһҗлҘј 비нҷңм„ұнҷ” мӢңмј°мқ„ л•Ң к°„мЎ°м§Ғм—җ мӨ‘м„ұм§Җл°©мқҳ 축м Ғмқҙ м•Ҫ 50% м •лҸ„ к°җмҶҢн•Ёмқ„ ліҙкі н•ң м—°кө¬лҘј нҶөн•ҙ Srebf1к°Җ мқёмҠҗлҰ° м Җн•ӯм„ұлҸҷл¬ј лӘЁлҚём—җм„ң м§Җл°©к°„мқҳ л°ңмғқм—җ мӨ‘мҡ”н•ң м—ӯн• мқ„ лӢҙлӢ№н•Ёмқ„ м•Ң мҲҳ мһҲлӢӨ[16].
мң лҰ¬м§Җл°©мӮ°мқҖ лІ нғҖ мӮ°нҷ”лҘј нҶөн•ҳм—¬ м—җл„Ҳм§ҖмӣҗмңјлЎң м „нҷҳлҗҳм–ҙм•ј н•ҳм§Җл§Ң ліё м—°кө¬мқҳ кІ°кіјмҷҖ к°ҷмқҙ м§ҖлӮҳм№ң м§Җл°©мқҳ 축м ҒмқҖ лІ нғҖмӮ°нҷ”лҘј м–өм ңн•ҳкі (Fig. 2E) Cyp2e1мқҳ л°ңнҳ„мқ„ мҰқк°ҖмӢңмјң лҢҖмІҙ мӮ°нҷ”кіјм •мқ„ мқјмңјнӮӨкІҢ н•ңлӢӨ(Fig. 2F). к°„м„ёнҸ¬мқҳ м§Җл°©мӮ° мӮ°нҷ”нҡЁмҶҢмқё Cyp2e1мқҖ нҷңм„ұмқҙ мҰқк°Җн•ҳл©ҙ к°„м—җ 축м Ғлҗң мң лҰ¬м§Җл°©мӮ°мңјлЎңл¶Җн„° нҷңм„ұмӮ°мҶҢмў…мқ„ мғқм„ұмӢңмјң мӮ°нҷ”мҠӨнҠёл ҲмҠӨлҘј мқјмңјнӮӨлҠ” кІғмңјлЎң м•Ңл Өм ёмһҲлӢӨ[17]. м„ёнҸ¬мқҳ лҢҖмӮ¬кіјм •м—җм„ң лҜёнҶ мҪҳл“ңлҰ¬м•„лҠ” ATPлҘј мғқм„ұн•ҳлҠ” кё°лҠҘмқ„ лӢҙлӢ№н•ңлӢӨ. мқҙ кіјм • мӨ‘м—җ лҜёнҶ мҪҳл“ңлҰ¬м•„лҠ” мғқмЎҙм Ғ л°ҳмқ‘мңјлЎң мӮ°мҶҢлЎң м „мһҗмқҳ мқҙлҸҷмқҙ мҰқк°Җлҗҳм–ҙ нҷңм„ұмӮ°мҶҢмў…(reactive oxygen species, ROS)мқҙ мғқм„ұн•ҳлҠ”лҚ°, м§ҖлӮҳм№ң ROSлҠ” IL-6мҷҖ TNFлҘј нҸ¬н•Ён•ң м—¬лҹ¬ мӮ¬мқҙнҶ м№ҙмқёмқҳ м „мӮ¬мқёмһҗмқё NF-kBлҘј нҷңм„ұнҷ”мӢңмјң лҜёнҶ мҪҳл“ңлҰ¬м•„мқҳ кё°лҠҘм Җн•ҙмҷҖ ROS мғқм„ұмқ„ мҰқк°ҖмӢңнӮӨкІҢ лҗңлӢӨ. лҜёнҶ мҪҳл“ңлҰ¬м•„ л§үм—җ мЎҙмһ¬н•ҳлҠ” UCP2лҠ” мқҙлҹ¬н•ң ROSмқҳ мғқм„ұмқ„ м–өм ңн•ҳлҠ” лӢЁл°ұм§ҲлЎң 비м•ҢмҪңм„ұ м§Җл°©к°„м§Ҳнҷҳ (non-alcoholic fatty liver disease, NAFLD)кіј 비л§Ңм—җм„ң мӮ°нҷ”мҠӨнҠёл ҲмҠӨ(oxidative stress)к°Җ лҶ’мқ„ л•Ң л°ңнҳ„мқҙ мҰқк°Җн•ңлӢӨкі ліҙкі лҗҳм–ҙмһҲлӢӨ[18]. ліё м—°кө¬м—җм„ңлҸ„ 16мЈјлҸҷм•Ҳ 45%мқҳ кі м§Җл°©мӢқмқҙлҘј м Ғмҡ©н•ң мғқмҘҗмқҳ м§Җл°©мЎ°м§Ғм—җм„ң Ucp2мҷҖ мӮ¬мқҙнҶ м№ҙмқё(IL6, Tnf)мқҳ мҰқк°ҖлҘј нҷ•мқён• мҲҳ мһҲм—ҲлҠ”лҚ°(Fig. 3C), мқҙлҠ” м§Җл°©мӮ°м—җм„ң мғқм„ұлҗң нҷңм„ұмӮ°мҶҢмқҳ мҰқк°Җм—җ л”°лқј мҰқк°Җн•ң мқјл Ёмқҳ л°ҳмқ‘мқҙлқј ліј мҲҳ мһҲлӢӨ.
ліё м—°кө¬м—җм„ң Lep, Nr3c1 к·ёлҰ¬кі Hsd11b1 м„ё к°Җм§Җ мң м „мһҗлҠ” мӢқмқҙмқҳ кі м§Җл°© н•Ёлҹүм—җ л”°лқј л°ңнҳ„мқҳ м°ЁмқҙлҘј ліҙмҳҖлӢӨ. LepмқҖ м§Җл°©м„ёнҸ¬м—җм„ң 분비лҗҳлҠ” нҳёлҘҙлӘ¬ мң м „мһҗлЎң м •мғҒмӢқмқҙлҘј н•ҳлҠ” л§Ҳмҡ°мҠӨм—җ 비н•ҙ 45HFDкө°м—җм„ң м•Ҫ 6л°°, 60HFDкө°м—җм„ңлҠ” м•Ҫ 21.5л°°мқҳ mRNA л°ңнҳ„мқҳ мҰқк°ҖлҘј ліҙмқёлҚ° 비н•ҙ мӢқмқҙ м„ӯм·Ёлҹүм—җлҠ” нҒ° ліҖнҷ”лҘј ліҙмқҙм§Җ м•ҠмқҖ кІғмңјлЎң ліҙм•„ м§Җл°©лҹүмқҳ мҰқк°ҖлҘј к°„м ‘м ҒмңјлЎң лӮҳнғҖлӮҙкё°лҸ„ н•ҳл©°[19, 20], 비л§Ңкө°м—җм„ң нқ”нһҲ лӮҳнғҖлӮҳлҠ” л ҷнӢҙ м Җн•ӯм„ұмқҳ кІ°кіјлЎңлҸ„ н•ҙм„қн• мҲҳ мһҲлӢӨ. 60HFDкө°м—җм„ң мӮ¬лЈҢм„ӯм·Ёлҹүмқҙ м•Ҫ 20% к°җмҶҢн•ҳмҳҖмңјлӮҳ мӢӨм ң л ҷнӢҙмқҳ м Җн•ӯм„ұмқҙ мһҲлҠ”м§ҖлҠ” л ҷнӢҙ мҲҳмҡ©мІҙлҘј нҸ¬н•Ён•ҳлҠ” кҙҖл Ё нҳёлҘҙлӘ¬мқҳ л°ңнҳ„кіј 분비м—җ лҢҖн•ң нӣ„мҶҚ мЎ°мӮ¬к°Җ н•„мҡ”н• кІғмңјлЎң ліҙмқёлӢӨ. лҳҗн•ң, кёҖлЈЁмҪ”мҪ”лҘҙнӢ°мҪ”мқҙл“ңлҠ” 비л§Ңкіј 비м•ҢмҪңм„ұ м§Җл°©к°„м§Ҳнҷҳм—җ м—°кҙҖм„ұмқҙ мһҲлҠ” кІғмңјлЎң м•Ңл ӨмЎҢлӢӨ. Andrews л“ұмқҳ м—°кө¬м—җ мқҳн•ҳл©ҙ л¶Ҳнҷңм„ұнҳ• мҪ”лҘҙнӢ°мҶҗ(cortisone)мқ„ нҷңм„ұнҳ• мҪ”нӢ°мҶ”(cortisol)лЎң ліҖнҷҳмӢңнӮӨлҠ” нҡЁмҶҢмқё Hsd11b1лҘј м–өм ңн•ҳл©ҙ мІҙмӨ‘ л°Ҹ нҳҲмӨ‘ м§Җм§ҲмҲҳм№ҳмқҳ к°җмҶҢнҷ” н•Ёк»ҳ к°„мқҳ мқёмҠҗлҰ° м Җн•ӯм„ұмқҙ лӮ®м•„진лӢӨкі ліҙкі н•ҳмҳҖлӢӨ[21]. мқҙлҠ” ліё м—°кө¬кІ°кіјм—җм„ңлҸ„ мң мӮ¬н•ҳкІҢ лӮҳнғҖлӮҳ кі м§Җл°©мӢқмқҙлҘј н• мҲҳлЎқ м§Җл°©м„ёнҸ¬мқҳ Hsd11b1мқҳ л°ңнҳ„кіј н•Ёк»ҳ мҲҳмҡ©мІҙмқё Nr3c1 мқҳ л°ңнҳ„лҸ„ мҰқк°Җн•ҳлҠ” кІғмқ„ ліј мҲҳ мһҲм—ҲлӢӨ. к·ёлҹ¬лӮҳ к°„мЎ°м§Ғм—җм„ң Hsd11b1мқҳ л°ңнҳ„мқҖ нҳ„м Җн•ҳкІҢ к°җмҶҢн•ҳкі (Fig. 3H), нҳҲмӨ‘ мҪ”лҘҙнӢ°мҪ”мҠӨн…ҢлЎ мқҳ мҲҳм№ҳм—җлҠ” ліҖнҷ”к°Җ м—ҶлҠ” кІғмңјлЎң ліј л•Ң, м „мӢ м Ғмқё нҳҲм•Ў лӮҙ кёҖлЈЁмҪ”мҪ”лҘҙнӢ°мҪ”мқҙл“ңмҷҖ мЎ°м§Ғм—җм„ң л°ңнҳ„лҗҳлҠ” нҳёлҘҙлӘ¬мқҳ м–‘мқҖ лҸҷмқјн•ҳм§Җ м•ҠлӢӨлҠ” м җмқ„ м•Ң мҲҳ мһҲлӢӨ. лҳҗн•ң к°Ғ мЎ°м§Ғл§ҲлӢӨ л°ңнҳ„мқҳ м •лҸ„к°Җ лӢӨлҘҙлҜҖлЎң мқҙ нҳёлҘҙлӘ¬м—җ мқҳн•ң лҢҖмӮ¬кё°лҠҘмқҳ мЎ°м Ҳм—җ лҢҖн•ҙ м•һмңјлЎң лҚ” м—°кө¬лҗҳм–ҙм•ј н• кІғмқҙлӢӨ. лҳҗн•ң, мқёмҠҗлҰ°м Җн•ӯм„ұмқҖ 비л§ҢмқҙлӮҳ м ң2нҳ•лӢ№лҮЁмқҳ м Җл“ұкёү м—јмҰқ(low-grade inflammation) мғҒнғңм—җ мқҳн•ҙ лҸҷл°ҳлҗҳм–ҙ м—јмҰқ мӮ¬мқҙнҶ м№ҙмқёмқ„ 분비мӢңнӮӨкі , 11ОІHSD1мқ„ мҰқк°ҖмӢңмјң кёҖлЈЁмҪ”мҪ”лҘҙнӢ°мҪ”мқҙл“ңлҘј нҷңм„ұнҷ”мӢңнӮЁлӢӨ[22]. TNF лҳҗлҠ” dexamethasoneмқҙ мҘҗмқҳ м§Җл°©м„ёнҸ¬м—җм„ң мқёмҠҗлҰ°м Җн•ӯм„ұмқ„ мң лҸ„н•ңлӢӨкі ліҙкі лҗҳм—ҲлӢӨ[23]. Foxo1мқҖ мқёмҠҗлҰ° мӢ нҳём „лӢ¬мқ„ л§Өк°ңн•ҳлҠ” м „мӮ¬мЎ°м ҲмқёмһҗлЎң мқёмҠҗлҰ° мҲҳмҡ©мІҙлҘј м ңкұ°н•ң мң м „мһҗліҖнҳ• мҘҗмқҳ к°„м—җм„ң мҰқк°Җлҗҳм—Ҳкі , Foxo1мқ„ м ңкұ°н•ҳл©ҙ лҜёнҶ мҪҳл“ңлҰ¬м•„мқҳ мӮ°нҷ” мӢ 진лҢҖмӮ¬мҷҖ мғқн•©м„ұ кё°лҠҘмқ„ нҡҢліөмӢңмј°лӢӨ[24]. ліё м—°кө¬ кІ°кіјм—җм„ң лӮҳнғҖлӮң Foxo1мқҳ мҰқк°ҖлҠ” лӢ№лҢҖмӮ¬мҷҖлҠ” мғҒкҙҖм—Ҷмқҙ лҜёнҶ мҪҳл“ңлҰ¬м•„мқҳ кё°лҠҘмқ„ лӮ®м¶”лҠ” кІғмңјлЎң ліҙмқёлӢӨ.
ліё м—°кө¬м—җм„ң кі м§Җл°©мӢқмқҙлҘј кіөкёүн•ң л§Ҳмҡ°мҠӨл“Өмқҳ м§Җл°©кіј к°„ мЎ°м§Ғмқҳ мғқмІҙмӢңкі„ мң м „мһҗл“Өмқҳ л°ңнҳ„мқҖ м§Җл°© н•Ёлҹүм—җ л”°лқј мҰқк°Җн•ҳлҠ” нҢЁн„ҙмқ„ ліҙмҳҖлӢӨ. лҳҗн•ң 16мЈјл № 45HFDкө°мқҳ нҳҲмӨ‘ мғқнҷ”н•ҷ м§Җн‘ңк°Җ 8мЈјл № 60HFDкө°мқҳ м§Җн‘ңл“Өл§ҢнҒј мғҒмҠ№н•ҳмҳҖлҠ”лҚ° мғқмІҙмӢңкі„ мң м „мһҗл“Өмқҳ л°ңнҳ„ м •лҸ„к°Җ лҸҷмқјн•ҳкІҢ мғҒмҠ№лҗҳлҠ” кІғмқ„ нҷ•мқён• мҲҳ мһҲм—ҲлӢӨ. лӢЁ, 60HFDкө°мқҳ к°„мЎ°м§Ғкіј 16мЈјл № 45HFDкө° м§Җл°©мЎ°м§Ғмқҳ Per2, 3мқҳ мң м „мһҗ л°ңнҳ„мқҙ нҳ„м Җн•ҳкІҢ к°җмҶҢн•ҳлҠ” кІғмңјлЎң ліҙм•„ мң м „мһҗл“Өмқҳ м•„мқҙмҶҢнҳ•(isoform)л“Өмқҙ к°Ғк°Ғ лӢӨлҘё кё°лҠҘмқ„ к°Җм§Җкі мһҲлҠ” кІғмңјлЎң ліҙмқҙл©° мЎ°м§Ғм—җ л”°лқј мң м „мһҗмқҳ л°ңнҳ„нҢЁн„ҙкіј мЎ°м Ҳмқёмһҗк°Җ лӢӨлҘј кІғмқ„ мҳҲмғҒн• мҲҳ мһҲлӢӨ. м§Җл°©м„ёнҸ¬мқҳ мЈјмҡ” м—…л¬ҙлҠ” м§Җл°©лҢҖмӮ¬мҷҖ м—јмҰқ мӮ¬мқҙнҶ м№ҙмқёмқҳ 분비мқҙл©°, Clockкіј ArntlмқҖ кі м§Җл°©мӢқмқҙм—җ мқҳн•ң м§Җл°©лҢҖмӮ¬мқҳ нҷңм„ұмқ„ мЎ°м Ҳн•ҳлҠ” кІғмңјлЎң мғқк°ҒлҗңлӢӨ[25]. Clockкіј Arntlмқҳ нҷңм„ұмңјлЎң мҰқк°Җлҗң CryмҷҖ Per л°ңнҳ„мқҳ мҰқк°ҖлҠ” negative feedbackм—җ мқҳн•ҳм—¬ Clockкіј Arntlмқҳ л°ңнҳ„мқ„ м–өм ңн•ҳлҠ” нҢЁн„ҙмқ„ ліҙмҳҖлӢӨ.
кі м§Җл°©мӢқмқҙлЎң мң лҸ„н•ҳлҠ” 비л§Ңм—°кө¬лҘј н• л•Ң 60%мқҳ кі м§Җл°© мӢқмқҙлҠ” 짧мқҖ мӢңк°„ лӮҙм—җ 비л§Ңмқ„ мң лҸ„н• мҲҳ мһҲм–ҙ мқјл°ҳм ҒмңјлЎң м“°мқҙлҠ” л°©лІ•мқҙлӮҳ 1 g лӢ№ 5.24 kcalлЎң 232 mgмқҳ мҪңл ҲмҠӨн…ҢлЎӨмқ„ н•Ёмң н•ҳкі мһҲм–ҙ мӮ¬лһҢмқҳ мӢқмқҙм—җ кІ¬мЈјм–ҙ ліҙл©ҙ нҳ„мӢӨм Ғмқҙлқј ліҙкё° м–ҙл өкі [9] 비л§Ң л°ңмғқмқҳ м •лҸ„к°Җ мқҙлҜё н•ңкі„м җмқ„ лҸ„лӢ¬н•ң мғҒнғңмқҙлҜҖлЎң ліё м—°кө¬м—җм„ңлҠ” 45% кі м§Җл°©мӢқмқҙлЎң мӨ‘к°„ м •лҸ„мқҳ 비л§Ңмқ„ мң лҸ„н•ҳкі мқҙлҘј мҠӨнҠёл ҲмҠӨк°Җ к°ҖмҶҚмӢңнӮ¬ мҲҳ мһҲлҠ”м§Җ ліҙкі мһҗ н•ҳмҳҖлӢӨ. л¶ҖлҸҷ мҠӨнҠёл ҲмҠӨ(immobilization stress) лӘЁлҚёмқҖ л§Ңм„ұ мҠӨнҠёл ҲмҠӨлҘј м Ғмҡ©н•ҳлҠ”лҚ° мһҗмЈј м“°мқҙлҠ” л°©лІ•мңјлЎң мӢңмғҒн•ҳл¶Җ-лҮҢн•ҳмҲҳмІҙ-л¶ҖмӢ 축(hypothalamus-pituitary-adrenal axis)кіј көҗк°җмӢ кІҪкі„(sympathetic nervous system)мқҳ нҷңм„ұнҷ”лҘј нҠ№м§•мңјлЎң н•ңлӢӨ. Thaker л“ұмқҳ м—°кө¬м—җ мқҳн•ҳл©ҙ нҠ№мҲҳ м ңмһ‘лҗң м•„нҒ¬лҰҙ мјҖмқҙмҠӨм—җ л§Өмқј 2мӢңк°„м”© к°җкёҲлҗң л§Ҳмҡ°мҠӨл“Өмқҳ мҠӨнҠёл ҲмҠӨ нҳёлҘҙлӘ¬мқё нҳҲмӨ‘ мҪ”лҘҙнӢ°мҪ”мҠӨн…ҢлЎ мқҖ мӢӨн—ҳ к°ңмӢң 3мқјл¶Җн„° мҰқк°Җн•ҳкё° мӢңмһ‘н•ҳм—¬ 7мқјкіј 14мқјм—җлҠ” лҢҖмЎ°кө°м—җ 비көҗн•ҳм—¬ мң мқҳн•ҳкІҢ мҰқк°Җн•ҳмҳҖлӢӨ. к·ёлҹ¬лӮҳ, мһҘкё°к°„мқҳ мҠӨнҠёл ҲмҠӨ м Ғмҡ©мқҙ нҳҲмӨ‘ мҪ”лҘҙнӢ°мҪ”мҠӨн…ҢлЎ мқ„ м§ҖмҶҚм ҒмңјлЎң лҶ’мқҖ мғҒнғңлЎң мң м§Җн•ҳм§ҖлҠ” м•ҠлҠ” кІғмңјлЎң ліҙмқёлӢӨ[10]. Voorheesмқҳ м—°кө¬м—җ л”°лҘҙл©ҙ, н•ҳлЈЁ 6мӢңк°„м”© л¶ҖлҸҷ мҠӨнҠёл ҲмҠӨлҘј м Ғмҡ©н•ҳмҳҖмқ„ л•Ң м Ғмҡ© мҙҲкё°мқё 7мқјм—җ к°ҖмһҘ мҲҳм№ҳк°Җ лҶ’м•ҳмңјлӮҳ мқҙнӣ„ м җм°Ё к°җмҶҢн•ҳлҠ” 추세лҘј ліҙмҳҖлӢӨ[26]. м§ҖмҶҚм Ғмқё мҠӨнҠёл ҲмҠӨк°Җ мһҘкё°м ҒмңјлЎң м Ғмҡ©лҗҳм—ҲмқҢм—җлҸ„ л¶Ҳкө¬н•ҳкі мҠӨнҠёл ҲмҠӨ нҳёлҘҙлӘ¬мқҙ к°җмҶҢн•ҳлҠ” мқҙмң мҷҖ кё°м „м—җ лҢҖн•ҙм„ңлҠ” м•Ңл Ө진 л°”к°Җ м—ҶмңјлӮҳ л‘җ к°Җм§Җмқҳ кІҪмҡ°лЎң м„ӨлӘ…мқҙ к°ҖлҠҘн• кІғмңјлЎң ліҙмқёлӢӨ. мІ«м§ё, м •мғҒм Ғмқё мҠӨнҠёл ҲмҠӨ нҳёлҘҙлӘ¬мқҖ лӮ®кіј л°Өмқҳ мқјмЈјкё°м—җ л”°лқј мҲҳм№ҳмқҳ кі м Җк°Җ мһҲлҠ” мқјліҖнҷ”(diurnal cycle)лҘј к°Җм§ҖлҠ” л°ҳл©ҙ, мһҘкё°м Ғ мҠӨнҠёл ҲмҠӨк°Җ м Ғмҡ©лҗЁм—җ л”°лқј мқјліҖнҷ”мқҳ мЈјкё°к°Җ лӢЁм¶• лҳҗлҠ” м§Җм—°мқҙ лҗ к°ҖлҠҘм„ұмқҙ мһҲлӢӨ. л‘ҳм§ё, мҠӨнҠёл ҲмҠӨ м Ғмҡ© мҙҲкё°м—җлҠ” мҰқк°Җн–ҲлҚҳ мҠӨнҠёл ҲмҠӨ нҳёлҘҙлӘ¬ мҲҳм№ҳк°Җ к°җкёҲмқ„ н•ҙм ңн•ҳлҠ” лҸҷмӢңм—җ кё°мӨҖм№ҳ(baseline)лЎң кёүкІ©нһҲ л–Ём–ҙм§ҖлҚҳ кІғмқҙ мҠӨнҠёл ҲмҠӨ м Ғмҡ©кё°к°„мқҙ кёём–ҙм§ҲмҲҳлЎқ 18мӢңк°„ мқҙмғҒ лҶ’мқҖ мҲҳм№ҳлҘј мң м§Җн•ҳлҠ” кІғмңјлЎң ліҙм•„[26], л§Ңм„ұ мҠӨнҠёл ҲмҠӨлҠ” нҳёлҘҙлӘ¬мқҳ мҲҳм№ҳмқҳ мқјліҖнҷ”к°Җ м—Ҷм–ҙм§Җл©ҙм„ң мқјм • мҲҳмӨҖмқ„ м§ҖмҶҚм ҒмңјлЎң мң м§Җн• кІғмңјлЎң ліҙмқёлӢӨ. к·ёлҹ¬лӮҳ ліё м—°кө¬м—җм„ңлҠ” мҪ”лҘҙнӢ°мҪ”мҠӨн…ҢлЎ мқ„ мӢңк°„ к°„кІ©мқ„ л‘җкі мЈјкё°м ҒмңјлЎң мёЎм •н•ҳм§Җ м•Ҡм•ҳкі 8мЈјмҷҖ 16мЈјм—җ к°Ғк°Ғ нҡЎлӢЁм ҒмңјлЎң мёЎм •н•ҳмҳҖкё° л•Ңл¬ём—җ нҳёлҘҙлӘ¬мқҳ мқјліҖнҷ” мЈјкё°м—җ ліҖнҷ”к°Җ мһҲм—ҲлӢӨл©ҙ мқҙлҘј лӢЁл©ҙ 비көҗн•ҳкё°м—җ м ңн•ңмқҙ мһҲм—Ҳмқ„ кІғмңјлЎң 여겨진лӢӨ.
м—¬лҹ¬ мһ„мғҒм—°кө¬м—җм„ң мҠӨнҠёл ҲмҠӨлҠ” 비л§Ңмқҳ мӣҗмқё мӨ‘ н•ҳлӮҳмқё кІғмңјлЎң мЈјлӘ©лҗҳм–ҙмҷ”лӢӨ. мІҙлӮҙ мҪ”нӢ°мҶ”мқҳ кіјлӢӨ분비м—җ мқҳн•ң мҝ мӢұмҰқнӣ„кө° нҷҳмһҗмқҳ мҰқмғҒм—җм„ңмҷҖ л§Ҳм°¬к°Җм§ҖлЎң мҪ”нӢ°мҶ”мқҳ мҰқк°ҖлҠ” нҸ¬лҸ„лӢ№мӢ н•©м„ұ(gluconeogenesis)мҷҖ м§Җм§Ҳ분н•ҙлҘј мң лҸ„н•ҳкі , л ҷнӢҙ м Җн•ӯм„ұмқ„ мҰқк°ҖмӢңмјң мІҙмӨ‘мқҳ мҰқк°ҖмҷҖ кі м—ҙлҹү мқҢмӢқм—җ лҚ” нғҗлӢүн•ҳкІҢ н•ҳлҠ” н–үлҸҷ ліҖнҷ”лҘј мқјмңјнӮЁлӢӨ[27]. к·ёлҹ¬лӮҳ мӮ¬лһҢмқҳ кІҪмҡ°мҷҖлҠ” лӢӨлҘҙкІҢ ліё м—°кө¬ кІ°кіјлҠ” мҠӨнҠёл ҲмҠӨ м Ғмҡ©мқҙ л ҷнӢҙ мҲҳм№ҳк°Җ мҰқк°Җн•ҳкі мӢқмқҙнҡЁмңЁмқҙ л–Ём–ҙмЎҢмқҢм—җлҸ„ л¶Ҳкө¬н•ҳкі мІҙмӨ‘кіј нҳҲмӨ‘ м§Җн‘ңл“Өмқҙ к°җмҶҢн•ҳлҠ” кІғмңјлЎң лӮҳнғҖлӮҳ мҠӨнҠёл ҲмҠӨмқҳ м Ғмҡ© л°©лІ•мқҙлӮҳ кё°к°„мқҙ м Ғн•©н•ҳм§Җ м•Ҡм•ҳмқ„ к°ҖлҠҘм„ұмқ„ ліҙм—¬мЈјкі мһҲлӢӨ. мқҙлҠ” м§Җл°© мЎ°м§Ғкіј к°„мқҳ лҢҖмӮ¬ кҙҖл Ё мң м „мһҗл“Өмқҳ мң мқҳн•ң ліҖнҷ”к°Җ ліҙмқҙм§Җ м•Ҡкі мҳӨнһҲл Ө м§Җл°©мӮ°мқҳ лІ нғҖ мӮ°нҷ”лҘј мҙү진н•ҳлҠ” кІ°кіјмҷҖ мқјм№ҳн•ҳкі мһҲлӢӨ. м•„мҡёлҹ¬ мҠӨнҠёл ҲмҠӨлҘј м Ғмҡ©н•ң мғқмҘҗл“Өмқҳ мғқмІҙмӢңкі„ мң м „мһҗ л°ңнҳ„мқҙ 45% кі м§Җл°©мӢқмқҙлҘј мӨҖ мғқмҘҗл“Өмқҳ мҲҳм№ҳм—җ 비н•ҳм—¬ к°җмҶҢн•ң кІғмқҖ м§Җл°©н•©м„ұмқҙ мӨ„кі мҶҢлӘЁк°Җ лҠҳм–ҙлӮЁм—җ л”°лқј мғқмІҙм§Җн‘ңл“Өмқҙ мҳӨнһҲл Ө к°ңм„ лҗҳлҠ” кІғкіј м—°кҙҖм„ұмқҙ мһҲлҠ” кІғмңјлЎң ліҙмқёлӢӨ.
лӢ№лҢҖмӮ¬мҷҖ м§Җл°©лҢҖмӮ¬к°Җ л§җмҙҲ мғқмІҙмӢңкі„ мң м „мһҗмқҳ мЎ°м Ҳм—җ м–ҙл–Ө кё°м „мңјлЎң мһ‘мҡ©н•ҳлҠ”м§ҖлҠ” м•„м§Ғк№Ңм§Җ мһҳ м•Ңл Өм ёмһҲм§Җ м•ҠлӢӨ. л¬ҙм—ҮліҙлӢӨ лҢҖмӮ¬м—җ кҙҖм—¬н•ҳлҠ” мһҘкё°л“Өм—җ лӢЁмқј м„ёнҸ¬мҲҳмӨҖмқҳ л§җмҙҲ мғқмІҙмӢңкі„ мң м „мһҗл“Өмқҙ мЎҙмһ¬н•ҳл©° мҲҳл©ҙ-к°Ғм„ұ мЈјкё°мҷҖлҠ” лі„к°ңлЎң мЎ°м§Ғ л°Ҹ кё°кҙҖ лі„ нҠ№мқҙм Ғмқё мғқмІҙлҰ¬л“¬мқ„ мғқм„ұн• кІғмқҙлқјлҠ” к°ңл…җмқҙ мөңк·јм—җ лҢҖл‘җн•ҳмҳҖкё° л•Ңл¬ём—җ нҳ„мһ¬к№Ңм§ҖлҠ” мғқмІҙмӢңкі„ мң м „мһҗ мЎ°мһ‘мқ„ мқҙмҡ©н•ҳкұ°лӮҳ мҲҳл©ҙ-к°Ғм„ұ мЈјкё°лҘј көҗлһҖмӢңнӮЁ нӣ„ мғқлҰ¬м Ғ ліҖнҷ”лҘј кҙҖм°°н•ҳлҠ” м—°кө¬л“Өмқҙ лҢҖл¶Җ분мқ„ м°Ём§Җн•ҳкі мһҲлӢӨ. мқҙлҘј л’·л°ӣм№Ён•ҳлҠ” кё°м „мқҖ мғқмІҙмӢңкі„лҘј мЎ°м Ҳн•ҳлҠ” CLOCK, BMAL1мқҙ м „мӮ¬мқёмһҗ(transcription factor)мқҙл©° CRY, PER л“ұмқҙ мЎ°м Ҳмқ„ л°ӣлҠ” н•ҳмң„ мң м „мһҗл“ӨмқҙлҜҖлЎң м§Җл°©мғқм„ұм—җ кҙҖм—¬н•ҳлҠ” sterol regulatory element-binding protein (SREBP) лӮҳ CCAAT-enhancer-binding proteins (C/EBPs) л“ұмқҙ н‘ңм Ғмң м „мһҗлЎңм„ң мғқмІҙмӢңкі„мқҳ мЎ°м Ҳмқ„ л°ӣлҠ” кІғмңјлЎң м„ӨлӘ…н•ҳкі мһҲлӢӨ[28]. к·ёлҹ¬лӮҳ ліё м—°кө¬мІҳлҹј кі м§Җл°©мӢқмқҙ л“ұмқ„ нҶөн•ҙ 비л§Ңмқ„ мң лҸ„н•ҳмҳҖмқ„ л•Ңм—җлҸ„ мғқмІҙмӢңкі„ мң м „мһҗмқҳ ліҖнҷ”к°Җ мқјм–ҙлӮҳлҠ” м—°кө¬кІ°кіјлҸ„ мһҲм–ҙ[29] мғҒнҳём Ғмқё мЎ°м Ҳмқҙ мқјм–ҙлӮҳлҠ” кІғмңјлЎң ліҙмқёлӢӨ. м•„м§Ғк№Ңм§Җ л§җмҙҲ мғқмІҙмӢңкі„ мң м „мһҗмқҳ мғҒмң„ мӢ нҳём „лӢ¬мІҙкі„к°Җ нҷ•мӢӨнһҲ м•Ңл Ө진 кІғмқҙ м—ҶмңјлӮҳ мқјл¶Җ м—°кө¬м—җм„ң м—җл„Ҳм§Җ н•ӯмғҒм„ұмқ„ мЎ°м Ҳн•ҳлҠ” AMP-activated protein kinase (AMPK)мқҳ нҷңм„ұмқҙ PPARs кіј к°ҷмқҙ лҢҖмӮ¬м—җ кҙҖм—¬н•ҳлҠ” мЈјмҡ” м „мӮ¬мқёмһҗмқҳ нҷңм„ұмқ„ мң лҸ„н•ҳкі мқҙл“Өмқҳ н‘ңм Ғ мң м „мһҗлЎң мғқмІҙмӢңкі„ мң м „мһҗл“Өмқҳ л°ңнҳ„мқҙ мқјм–ҙлӮҳлҠ” кІғмңјлЎң ліҙкі лҗҳкі мһҲлӢӨ[6]. к·ёлҹ¬лӮҳ мқҙлҘј м§Җм§Җн•ҳлҠ” м—°кө¬л“Өмқҙ мҶҢмҲҳмқҙлҜҖлЎң м •нҷ•н•ң кё°м „мқ„ к·ңлӘ…н•ҳкё° мң„н•ң 추к°Җм Ғмқё м—°кө¬к°Җ н•„мҡ”н• кІғмқҙлӢӨ.
кІ°лЎ
ліё м—°кө¬лҠ” 45%мҷҖ 60%мқҳ кі м§Җл°©мӢқмқҙлҘј м„ӯм·Ён•ң л§Ҳмҡ°мҠӨ лӘЁлҚёмқ„ мқҙмҡ©н•ҳм—¬ м§Җл°© н•Ёмң лҹүкіј мӢқмқҙ кё°к°„м—җ л”°лҘё л§Ҳмҡ°мҠӨмқҳ мІҙмӨ‘, м„ӯмӢқ, нҳҲмӨ‘ мғқнҷ”н•ҷ м§Җн‘ңмҷҖ н•Ёк»ҳ лҢҖмӮ¬мҷҖ кҙҖл Ёлҗң мң м „мһҗ л°ңнҳ„мқҙ мҰқк°Җн•ҳкі м§Җл°©, к°„ мЎ°м§Ғмқҳ л§җмҙҲ мғқмІҙмӢңкі„ мң м „мһҗ л°ңнҳ„мқ„ мң лҸ„н•ҳлҠ” кІғмқ„ нҷ•мқён•ҳмҳҖлӢӨ. 45%мқҳ кі м§Җл°©мӢқмқҙлҘј м„ӯм·Ён•ң л§Ҳмҡ°мҠӨм—җкІҢ 16мЈјк°„ мһҘкё°м ҒмңјлЎң л¶ҖлҸҷ мҠӨнҠёл ҲмҠӨлҘј м Ғмҡ©н•ң кІҪмҡ° мҠӨнҠёл ҲмҠӨ нҳёлҘҙлӘ¬мқҳ ліҖнҷ”лҠ” мң мқҳн•ҳм§Җ м•Ҡм•ҳмңјл©° м§Җл°©мЎ°м§Ғмқҳ м§Җл°©н•©м„ұ мң м „мһҗк°Җ к°җмҶҢн•ҳкі к°„мЎ°м§Ғмқҳ м§Җл°©мӮ° лІ нғҖ мӮ°нҷ” нҡЁмҶҢ мң м „мһҗ л°ңнҳ„мқҙ мҰқк°Җн•ҳл©ҙм„ң нҳҲлӢ№кіј нҳҲмӨ‘ м§Җм§Ҳ мҲҳм№ҳк°Җ к°җмҶҢн•ҳлҠ” кІ°кіјлҘј ліҙмҳҖлӢӨ. кі м§Җл°©мӢқмқҙмҷҖ л¶ҖлҸҷ мҠӨнҠёл ҲмҠӨмқҳ м Ғмҡ©мқҖ л§җмҙҲ мЎ°м§Ғмқҳ мғқмІҙмӢңкі„ мң м „мһҗмқҳ л°ңнҳ„мқ„ ліҖнҷ”мӢңнӮӨлҜҖлЎң 추нӣ„ мғқмІҙмӢңкі„м—җ мқҳн•ң лҢҖмӮ¬мһ‘мҡ©мқҳ мЎ°м Ҳмқҳ мқҙн•ҙлҠ” 비л§Ң кҙҖл Ё м§Ҳлі‘мқҳ л°ңмғқмқҙлӮҳ мҲҳл©ҙ м–‘мғҒмқҙлӮҳ мӢқмқҙмқҳ ліҖнҷ”лҘј мң л°ңн•ҳлҠ” н–үлҸҷм—җ лҢҖн•ң мһ„мғҒм—°кө¬м—җ нҷңмҡ©лҗ мҲҳ мһҲмқ„ кІғмқҙлӢӨ.
 PDF Links
PDF Links PubReader
PubReader Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print